12. Конкуренция - реальные нейроны
Говоря о проблемах искусственного интеллекта, мы, естественно, пытаемся сопоставить полученные результаты с работой мозга, основанной на активности нейронов. Как правило, при таких сопоставлениях искусственный интеллект во многом проигрывает мозгу, о чем мы более подробно расскажем в последних трех главах нашей книги. Разумеется, превосходство мозга вполне вытекает из самого характера задач, которые ставятся в области искусственного интеллекта: эти задачи выбираются так, чтобы они были трудными для машин, но вполне доступными для мозга человека.
Машины, однако, превосходят человека в решении задач, которые требуют больших вычислительных работ, например исчерпывающего перебора вариантов. Если вычислительную машину удастся запрограммировать так, чтобы она была практически непобедимой в шахматах (что выглядит достаточно правдоподобным), то это наверняка будет достигнуто не имитацией методов игры человека, а посредством использования громадных объемов вычислительных ресурсов машины.
Хотя главная тема настоящей книги - искусственный интеллект, мы считаем тем не менее целесообразным уделить внимание структуре и функциям реальных нейронов, которые неразрывно связаны с естественным интеллектом. Некоторые аспекты функционирования систем нейронов представляют здесь особый интерес вследствие определенного подобия их и искусственных систем.
Размеры нервных клеток ничтожны: основное "тело" клетки имеет в поперечнике 30 мкм (микрон, 1 мкм = 10-6 м). У тела нервной клетки обычно имеется много отростков. Многие из них представляют собой ветвящиеся структуры, называемые дендритами, они простираются на 200-300 мкм от тела клетки. Большинство нейронов имеют также более длинный отросток, называемый аксоном, который может иметь протяженность от 50 мкм до нескольких метров. Аксон часто также имеет много разветвлений.
Нервные стволы (как мы просто говорим, "нервы") состоят из пучка нервных волокон, окруженных защитной оболочкой, наподобие подземного телефонного кабеля. Эти волокна, по существу, являются аксонами нервных клеток, переносящими нервные импульсы.
Вся нервная клетка, включая дендриты и аксоны, защищена мембраной, вследствие чего химический состав протоплазмы внутри клетки отличен от химического состава жидкой среды организма вне ее. Основное различие состоит в том, что протоплазма клетки богата калием, тогда как внешняя среда - натрием. Это различие поддерживается благодаря определенному свойству мембраны, известному под названием "натриевый насос", и приводит к наличию на мембране электрической разности потенциалов. Потенциал внутри покоящегося нейрона по отношению к внешней среде составляет примерно - 70 мВ.
Если какая-то небольшая область мембраны деполяризуется, т. е. указанная разность потенциалов между клеткой и окружением снимается, то в этой области мембраны возникает ток, который воздействует на соседние области, вызывая также и их деполяризацию. Затем примерно в течение миллисекунды мембрана возвращается в нормальное состояние. В целом, как только деполяризуется какая-то малая область клетки, в ней возникает "волна деполяризации", которая распространяется по аксону. Это и есть нервный импульс, который обеспечивает связь в нервной системе.
Некоторые нервные волокна (так называемые мякотные нервные клетки) покрыты оболочкой из вещества, называемого миелином, которое выполняет роль изолятора. Чем толще волокно, тем быстрее оно проводит. Однако при одном и том же размере волокна импульс быстрее распространяется в волокнах, покрытых миелином. (Это, видимо, обусловлено тем, что в волокнах, покрытых миелином, через каждые несколько миллиметров имеются так называемые "перехваты Ранвье", между которыми нервный импульс как бы "проскакивает".) На самом деле скорость распространения нервного импульса много ниже скорости передачи электрических сигналов по кабелю. В мякотном волокне она составляет обычно несколько десятков метров в секунду (максимум 100 м/с). Для небольших волокон зрительного нерва лягушки, не покрытых миелином, скорость распространения импульса, как показали измерения, не превышает нескольких десятков сантиметров в секунду.
Аксон - это отросток нейрона, проводящий нервные импульсы от тела клетки к иннервированным органам и другим нейронам. На теле нервной клетки и на ее дендритах имеются области (до 10 тыс. точек на один нейрон), в которых на клетку оказывают воздействие аксоны других нейронов. Эти области называются синапсами. Существуют возбуждающие синапсы (стимулирующие деполяризацию нейрона) и тормозные (затрудняющие возбуждение, т. е. деполяризацию нейрона вследствие воздействия других нейронов через возбуждающие синапсы). Передача нервного импульса через синапс носит химический характер, причем в разных синапсах встречаются различные передающие вещества (медиаторы).
Под внешним воздействием нейрон может "сработать", создавая выходной импульс, который далее распространяется по аксону. Обычно такое "срабатывание" происходит под воздействием импульсов, поступающих из других нейронов через возбуждающие синапсы. Между моментом прихода импульса в данный нейрон и появлением сигнала на его выходе всегда имеется задержка - так называемая синаптическая задержка, - равная примерно 1 мс.
Таким образом, "вычислительные элементы" мозга действуют очень медленно по сравнению с элементами вычислительных машин, где скорости передачи сигналов обычно измеряются в наносекундах (1 нс = 10-9 с). Высокая "вычислительная эффективность" мозга достигается не за счет больших скоростей выполнения операций на нейронном уровне, а благодаря одновременному действию большого числа нейронов; в отличие от этого в вычислительных машинах все операции фактически производятся последовательно.
Хотя мозг и периферийная нервная система существенно отличаются от вычислительной машины по своей организации, некоторые аспекты их работы можно исследовать примерно так же, как это делается при изучении электронной схемы. При прохождении нервного импульса в нейроне возникает электрическое возбуждение, которое можно зарегистрировать с помощью миниатюрного электрода, помещенного вблизи тела нейрона или нервного волокна. Для этой цели были разработаны специальные микроэлектроды. Тщательно регулируя положение микроэлектрода с помощью микрометрического винта, можно замерять, усиливать и регистрировать сигналы от одиночного нейрона.
Микроэлектроды имеют различные конструкции. Самые тонкие из них изготовлены в виде микропипеток - они представляют собой сильно "оттянутые" стеклянные трубочки, заполненные проводящим раствором. Кончик некоторых из них достигает в диаметре ничтожной доли микрона и может проникать сквозь мембрану нейрона, не оказывая влияния на его работу. Соединив микроэлектрод с усилителем, имеющим большое входное сопротивление (оно необходимо потому, что сопротивление микроэлектрода может превышать 100 МОм), можно убедиться в наличии на мембране разности потенциалов в 70 мВ. Для других измерений используются менее тонкие электроды. Они не проникают в клетку, но, будучи тщательно установлены в нужном положении, позволяют регистрировать сигналы, относящиеся к одному нейрону. Такие микроэлектроды можно сделать в виде как микропипетки, так и стеклянной трубочки, в которую заключена тонкая платиновая проволочка. Еще один способ изготовления микроэлектродов связан с использованием вольфрамовой проволоки, заостренной методом электролитического травления и почти до самого кончика покрытой специальным термолаком.
Можно также возбуждать нейроны и инициировать импульс в любой точке аксона, подавая в нужном месте короткий импульс электрического тока (хотя, как мы уже отмечали, нормальное возбуждение через синапсы имеет химическую природу). Таким образом, мы получаем возможность "подключиться" к нервной системе животного как для введения в нее сигналов, так и для регистрации происходящих в ней процессов. (При экспериментах на нервной системе животное обычно подвергают легкой анестезии. Она должна быть легкой, поскольку анестезирующие вещества влияют именно на нервную систему. В тех случаях, когда ставится цель изучить какую-то область нервной системы (но не мозг), животное можно подвергнуть децеребрированию. При этом большую часть его мозга удаляют под анестезией, после чего анестезия прекращается. Практикуют также вживление микроэлектрода в мозг животного под анестезией, что позволяет производить нужные записи или стимуляцию нервных центров, когда животное находится в полном сознании и, судя по внешним признакам, не испытывает неудобств.)
Нервная система животных исследовалась весьма интенсивно. Однако мы не будем здесь пытаться делать обзор работ в области нейрофизиологии. Весьма интересную попытку такого рода предпринял в своей книге Зоммергофф [1]. И хотя успехи в области нейрофизиологии достигнуты немалые, многое остается еще в высшей степени загадочным. Ниже мы рассмотрим два вопроса, имеющие особое отношение к искусственному интеллекту, а именно: нейрофизиологию зрения и свойство нейронной пластичности.
Зрение лягушки
В результате изучения зрительных систем животных в настоящее время сформировалось достаточно последовательное представление о начальных этапах обработки зрительной информации. Глаз представляет собой, по существу, камеру с линзой, создающей изображение на сетчатке, которая состоит из более сотни миллионов светочувствительных клеток - фоторецепторов (у человека около 120 млн. палочек и примерно 7 млн. колбочек). В начале исследований зрительной системы обычно предполагалось, что выходные сигналы от фоторецепторов передаются по зрительному нерву в мозг для анализа примерно так же, как это сделал бы телевизионный инженер, если бы он не имел возможности сканировать изображение с целью сжатия сигнала для передачи его по одному широкополосному каналу.
Однако изображение не может быть передано в мозг точно в таком виде, как оно было получено, для этого в зрительном нерве недостаточно волокон. Поскольку каждый зрительный нерв человека содержит около миллиона волокон, для передачи изображения информацию необходимо сжать. В сетчатке существует множество соединений между первичными рецепторами и ганглиозными клетками, аксоны которых являются волокнами зрительного нерва. Указанное сжатие должно происходить в них.
Исследование зрительной системы лягушки, проведенное Леттвином и др. [2] (включая автора настоящей книги), показало, что сетчатка лягушки обрабатывает поступающий на нее световой сигнал таким образом, который едва ли пришел бы в голову телеинженеру. В сетчатке лягушки имеется примерно миллион первичных рецепторов и в каждом зрительном нерве - около полумиллиона волокон. Эти волокна, или нейроны*, как оказалось, принадлежат одному из четырех типов; нейроны каждого типа реагируют специальным способом, который как-то связан с образом жизни лягушки. Кроме того, эти нейроны передают изображение (как и предположил бы инженер-телевизионщик) с сетчатки на поля нейронов, расположенных в противоположных относительно каждого глаза зрительных долях мозга лягушки. Однако характер этих четырех типов нейронов таков, что, по-видимому, обработка зрительного образа началась уже на уровне сетчатки, при этом из него "извлекаются" признаки, важные для лягушки.
* (Здесь автор прибегает к распространенному у нейрофизиологов понятию регистрируемой единицы (unit), которое мы переводим здесь словом нейрон. - Прим. перев.)
Четыре типа нейронов таковы:
1. Детекторы длительного контраста. Каждый из них реагирует на зрительное поле диаметром два угловых градуса и не срабатывает при включении или выключении света в помещении. Если в это поле входит край предмета, который либо темнее, либо светлее фона и имеет четкую границу, то нейрон "срабатывает" (возникает поток импульсов); разряд продолжается до тех пор, пока край остается в поле зрения.
На реакцию таких нейронов очень мало влияют действительные уровни освещенности. Почти не наблюдалось различия в реакциях, когда уровни освещенности фона и края предмета были настолько малыми, что сам экспериментатор едва различал край.
2. Детекторы выпуклого края. Это самые интересные из описанных нейронов, называемые обычно "детекторами мухи". Они совершенно не реагируют ни на изменение общей освещенности, ни на прямой край предмета, перемещающийся по рецептивному полю. Они срабатывают лишь на вхождение в поле малого объекта (не более 3 угловых градусов в диаметре), более темного, чем фон.
Леттвин обнаружил также и другие удивительные свойства этих нейронов. Так, он заметил, что если темная точка движется вместе с (как-то помеченным) фоном, то эти нейроны ее "игнорируют". Реакция отсутствует и в том случае, если фон перемещается, а пятно остается неподвижным. Но так только пятно начинает двигаться, нейрон срабатывает.
Однако результаты, полученные Леттвином и его коллегами, вызвали критические замечания, в частности, Гейза и Джекобсона [3], по мнению которых столь простую интерпретацию реакций нейронов вряд ли можно считать надежной. Они предложили описывать эти нейроны, рассматривая некие концентрические области в зрительном поле. Такая точка зрения трудно согласуется с описанной Леттвином реакцией лягушки на небольшие тела, расположенные на помеченном фоне.
3. Детекторы движущегося края. Эти детекторы с рецептивным полем шириной примерно двенадцать градусов реагируют на движущийся край объекта независимо от того, представляет ли это движение наползание темного на светлое или наоборот. Реакция существенно зависит от скорости движения и очень слабо от протяженности края или яркости освещения. Эти нейроны реагируют, хотя и слабо, на включение и выключение света.
4. Детекторы затенения. Они имеют большое рецептивное поле - диаметром до пятнадцати градусов. Выходной сигнал с этих детекторов передается по мякотным волокнам зрительного нерва со скоростью, достигающей 10 м/с. Оки реагируют на внезапное потемнение, которое может быть вызвано, например, тенью какого-то хищника, угрожающего лягушке.
Детекторы края названных двух типов позволяют лягушке опознать хищника и потенциальную жертву, отличную от "мухи". Можно предположить, что "детекторы мухи" и "детекторы затенения" также самым непосредственным образом связаны с особенностями жизни лягушки.
Зрение млекопитающих
После успешных экспериментов Леттвина и его коллег, проведенных в Массачусетском технологическом институте (США), два исследователя, Хьюбель и Визель, из расположенного поблизости Гарвардского университета приступили к исследованиям с помощью микроэлектродов зрительной системы млекопитающих. Вначале они экспериментировали на кошках, как часто поступают в нейрофизиологических исследованиях, а затем перешли к обезьянам. (Конечно, с кошками экспериментировать проще, так как их много, но обезьяны больше похожи на человека.)
Результаты, полученные Хьюбелем и Визелем [4], сильно отличаются от описанных ранее результатов Леттвина. Данные Хьюбеля и Визеля наводят па мысль, что зрительный нерв в самом деле производит отображение зрительного образа сетчатки так, как это делалось бы в телевизионной системе, - последующий анализ изображений происходит в мозге и это несмотря на то, что у млекопитающего число первичных фоторецепторов еще более отличается от числа волокон в зрительном нерве, чем у лягушки.
Одной из причин расхождения между данными Леттвина, с одной стороны, и Хьюбеля и Визеля - с другой, является то, что последние отводили сигналы от части мозга, которую нельзя считать соответствующей зрительным долям лягушки. Они отводили сигналы от зрительной коры головного мозга (называемой также затылочной корой вследствие ее расположения сзади головы, или стриарной корой из-за ее слоистой структуры, или полями 17 и 18 коры, причем основные результаты были получены в поле 17).
Кора головного мозга высших животных - это тот орган, который, как считается, отвечает за мышление и другие "высшие функции" мозга. У лягушки она, по существу, отсутствует. Зрительными центрами у лягушки являются так называемые зрительные доли - самые крупные структуры ее мозга. В мозге млекопитающих зрительным долям лягушки соответствуют структуры, называемые левым и правым верхними холмиками; с ними и соединены зрительные нервы. У млекопитающих, однако, верхние холмики играют скорее вспомогательную роль в зрительном восприятии, так как они осуществляют управление мышцами, определяющими направление взгляда. Чтобы обеспечить правильное направление взгляда, верхние холмики должны выделять общие свойства зрительного образа, в частности его движения. Поэтому функция верхних холмиков у млекопитающих, возможно, весьма близка к функции зрительных долей лягушки.
Тот факт, что, как показывает эксперимент, зрительная система млекопитающих, по-видимому, действительно работает наподобие телевизионной камеры (если говорить об отображении в кору), выглядит менее удивительным, если учесть, что зрительная информация передается таким образом и по всей нервной системе. Высокая разрешающая способность сетчатки и нейронные сети, предназначенные для обработки информации на уровне сетчатки, вероятно, оказались бы излишними для коры, но скорее всего они играют, свою роль в сочетании с верхними холмиками.
Сигналы передаются в зрительные центры мезга по двум структурам (по одной с каждой стороны), называемым коленчатыми телами. Все реакции от их нейронов являются концентрическими, т. е. ненаправленными. Это значит, что реакция может возникнуть при действии пятна света в любом участке небольшой круглой области зрительного поля. Часто оказывается, что нейрон по-разному реагирует на пятно света внутри некоторого круга и на пятно света, которое оказывается вне этого круга, но остается в пределах большого круга, окружающего первый. Свет во внутренней области может возбуждать данный нейрон, а свет во внешней области - тормозить его реакцию. Эти две области могут иметь различную цветовую чувствительность.
Зрительное поле отображается в зрительные центры коры головного мозга. В этой области коры встречаются концентрические нейроны, подобные тем, что имеются в коленчатых телах, но обнаруживаются также нейроны трех других типов, которым Хьюбель и Визель дали следующие названия: простые (хотя они не столь просты, как концентрические нейроны), сложные и гиперсложные.
Каждый из нейронов названных трех типов связан с определенной ориентацией в зрительном поле, разной для различных нейронов. Для некоторых из простых нейронов в зрительном поле существует такая линия, что световые образы, находящиеся вблизи линии содной стороны от нее, возбуждают нейрон, а расположенные с другой - подавляют его активность. На рис. 38 показана такая линия, причем точки, в которых свет вызывает возбуждение, отмечены крестиком, а те, что создают тормозящий эффект, - кружком. Реакция нейрона такого типа максимальна, когда граница света и тени совпадает с указанной линией, причем граница проходит так, что точки, создающие торможение, находятся в темноте, а вызывающие возбуждение - освещены. Таким образом, этот простой нейрон действует как специализированный детектор края, находящегося в определенном положении в зрительном поле.
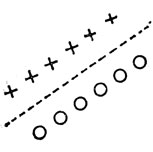
Рис. 38. Рецептивное поле нейрона того типа, который особенно сильно реагирует на присутствие края соответствующей ориентации. Крестики обозначают точки, в которых свет оказывает возбуждающее действие на нейрон, а кружочки - точки, где свет оказывает тормозящее действие
Хьюбель и Визель обнаружили и другие простые нейроны, способность которых к реакции может быть представлена так, как показано на рис. 39 и 40. Можно сказать, что они выделяют светлые и темные полоски соответственно. Интересно, что тип нейрона, соответствующий рис. 40, в точности совпадает с элементами, встроенными Парксом (в качестве этапа предварительной обработки) в его систему распознавания символов, предназначенных для обнаружения отрезков линий (гл. 7).
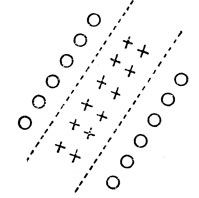
Рис. 39. Рецептивное поле нейрона, реагирующего на светлую полоску
Рис. 40. Рецептивное поле нейрона, реагирующего на темную полоску
Сложные нейроны, по существу, ведут себя так же, как простые, но реагируют только на движущиеся края и полоски. Хьюбель и Визель обнаружили их случайно, когда начали использовать края реальных объектов (типа листа картона) для того, чтобы вызвать быстрые реакции простых нейронов. Это, безусловно, более удобный способ, чем формирование изображений путем создания световых пятен последовательно по всей области. Исследователи обнаружили, что некоторые нейроны реагировали на край только в том случае, когда он двигался.
Сложные нейроны реагируют на движение края, а также светлых или темных полосок в определенной области зрительного поля, которую называют полем данного нейрона. На эти нейроны не действуют объекты, находящиеся вне их полей. Таким образом, не играет роли, простирается ли край или полоска за пределы этого поля или только соприкасается с ними. Но у гиперсложных нейронов имеется некое поле, в пределах которого движение края или полоски вызывает реакцию нейрона, и эта реакция максимальна, если объекты не выходят за пределы поля. При любом выходе объекта за пределы такого поля реакция нейрона уменьшается.
Довольно сложно понять, какой цели служат гиперсложные нейроны. Однако важно уже то, что один тип простого нейрона точно соответствует элементу, встроенному в искусственную систему. Что касается простого нейрона другого типа, то ему, по-видимому, также легко найти применение в искусственных схемах распознавания образов.
Реакция простого нейрона должна определяться комбинацией сигналов от некоторого числа концентрических нейронов. Аналогично сложный нейрон должен объединять выходы нескольких простых нейронов, имеющих одинаковую ориентацию. В таком случае гиперсложный нейрон должен объединять выходы нескольких сложных. Итак, получаем следующую прогрессию:
Не следует, однако, считать, что вся зрительная информация проходит по этому пути. Это следует хотя бы из того факта, что мы видим звезды в ночном небе. Здесь нет ни края, ни светлой или темной полоски, и тем не менее информация поступает в мозг. В экспериментах в области физиологии зрения было установлено, что на следующий этап обработки информации поступают входные данные от нейронов всех четырех типов. (Чтобы видеть звезды, сигнал должен поступать непосредственно от концентрических нейронов.) Нейроны указанных типов были обнаружены Хьюбелем и Визелем в поле 17. Следующий этап обработки зрительной информации, по-видимому, происходит в поле 18, но его работа пока плохо поддается описанию.
Хьюбель и Визель, подводя итог своим исследованиям, заявили следующее: "Специализированные нейроны, каковыми являются клетки поля 17, по сравнению с палочками и колбочками тем не менее должны представлять собой весьма элементарную стадию обработки сложных форм информации и заниматься, как это им свойственно, относительно простым анализом контуров областей, проецирующихся на сетчатку. То, как полученные при этом данные используются на последующих этапах обработки зрительной информации, представляет собой одну из наиболее интригующих проблем для будущих исследований".
Нейронная пластичность
Важная особенность естественного интеллекта заключается в его способности к обучению. Большинство видов активности мозга, которые мы пытаемся имитировать в искусственных системах, по крайней мере отчасти, являются результатом обучения на опыте. Многие исследователи считали, что путь к созданию настоящего искусственного интеллекта лежит через построение искусственных систем, способных обучаться. Примерами могут служить персептрон (гл. 8) и программа для игры в шашки Сэмюэля (гл. 6).
Процесс обучения в нервной системе, по-видимому, связан с определенными изменениями физических свойств нейронов (или, возможно, клеток другого типа, соседствующих с нейронами и влияющих на них). В персептроне и других модифицирующихся сетях реализуется гипотеза, согласно которой эти изменения сказываются на эффективности действия синапсов. Любая способность нейрона изменять свойства в зависимости от его предшествующей активности называется пластичностью. Большая часть экспериментальных работ (хотя, безусловно, далеко не все), касающихся пластичности, была выполнена на зрительной системе земноводных (лягушек и др.) или млекопитающих, о чем мы уже говорили.
Интересно узнать, например, связано ли появление простых, сложных и гиперсложных нейронов Хьюбеля и Визеля со зрительным опытом животного. Как это часто случается в биологических исследованиях, абсолютно четкого и ясного ответа на этот вопрос получить не удалось. Хьюбель и Визель обнаружили, что зрительный опыт животного не является необходимой предпосылкой для возникновения специализированных нейронов. Их развитие, однако, зависит от такого опыта. Одну группу котят специально содержали в таких условиях, что их опыт сводился лишь к восприятию вертикальных полос (большую часть времени они проводили в клетке в темноте, а когда внутренность клетки, выкрашенной вертикальными полосами, освещалась, их головы поддерживали в вертикальном положении). Другая группа подопытных котят имела дело лишь с горизонтальными полосами. Было установлено, что у первой группы котят почти все специализированные нейроны Хьюбеля и Визеля имеют вертикальную ориентацию, а у второй - горизонтальную. Поведение котят также вполне соответствовало результатам этих электрофизиологических исследований. Котята игриво бросались на любой предмет типа карандаша, которым помахивали перед ними. При этом котята первой группы не реагировали на горизонтально расположенный карандаш, и наоборот, котята второй группы "проявляли безразличие" к предметам, вытянутым по вертикали.
В последние годы ряд исследователей, прежде всего Уолл, занимались изучением другого класса явлений, связанных с пластичностью. Когда нейрон лишают входа, перерезая ведущие к нему пути, он часто начинает реагировать на другой вход, к которому прежде был нечувствителен. Такие эффекты изучались главным образом в связи с тактильными раздражителями.
В одной своей работе, которая, собственно, и привлекла внимание к этому эффекту, Уолл и Эггер [7] экспериментировали со взрослыми крысами, записывая сигналы из определенной области нижней части мозга, на которую проецировалась поверхность тела. Размеры этой области в поперечнике не превосходят нескольких миллиметров, но с помощью микроэлектродов исследователи смогли обнаружить на каждой стороне области отображения передней и задней конечности противоположной стороны животного. Эти области отображения можно было проследить, найдя хотя бы по одной точке в каждый момент времени. Кончик микроэлектрода, подсоединенного к усилителю, вводили в предполагаемую область отображения, а затем кожу крысы терли щеткой (со средним усилием) до тех пор, пока не наталкивались на точку, прикосновение к которой вызывало сигнал на усилителе.
У некоторых животных Уолл и Эггер прерывали путь, по которому сигналы от задней конечности шли к области отображения. В день такой операции можно убедиться, что существует "немая область" в том участке мозга, куда должна быть отображена задняя конечность. Через некоторое время область отображения передней конечности расширялась, как будто она была нарисована на резиновой пленке и теперь, вытянувшись, захватывала область, прежде занятую отображением задней конечности. Время, в течение которого происходили такие изменения, составляло две-три недели.
Одна из причин, по которой мог возникнуть эффект "резиновой пленки", состоит в физическом изменении нейронов в области отображения. Так могло бы случиться, если бы происходило сжатие неиспользуемых нейронов, которые представляли раньше заднюю конечность, и увеличение размеров остальных. Уолл и Эггер высказали соображения, позволяющие думать, что эффект "резиновой пленки" создается не таким образом, а обусловлен реальными изменениями в связях нейронов, в результате которых нейроны, прежде реагировавшие на стимуляцию какой-то точки на задней конечности, начинают теперь откликаться на возбуждение некоторой точки на передней конечности.
Изменение в связях может произойти в основном двумя путями: путем разрастания новых соединений либо путем демаскирования старых соединений, которые все время существовали, но не действовали. В настоящее время (конец 1981 г.) из этих двух возможностей Уолл считает наиболее вероятным демаскирование. Нейроны имеют множество соединений (часто до 10 тыс. на одну клетку), но наблюдаемые реакции можно объяснить с учетом, возможно, двух-трех входов. Поскольку избыточность во взаимных соединениях громадна, открываются широкие возможности для образования новых рисунков соединений путем демаскирования прежде не работавших синапсов.
Пластичность, исследованная Уоллом и его коллегами, обнаруживается лишь при хирургическом вмешательстве: важно действительное перерезание пути передачи сигналов в нейрон, а не просто прекращение подачи сигналов. Поэтому маловероятно, чтобы такого рода пластичность играла какую-то роль в обучении. С другой стороны, не ясно, каким образом синапс "узнает", что путь к нему где-то прерван. Недавно Уолл высказал предположение, что этот эффект, возможно, зависит от того факта, что при перерезании нервного пути происходит прерывание тонких не покрытых миелином волокон (так называемых С-волокон), переплетающихся с намного более крупными волокнами основного пути. Согласно некоторым данным (правда, пока недостаточно убедительным), С-волокна определяют эффективные соединения, сформированные в основных нервных путях. Если эта интерпретация верна, то, возможно, результаты Уолла имеют непосредственное отношение к процессу обучения.
Данные, полученные Уоллом и его коллегами, особенно интересны тем, что они касаются пластичности у взрослых животных в отличие от ранее упомянутых исследований на котятах. У молодых животных трудно отличить эффекты, которые скорее следует рассматривать как повзросление, от тех, которые вызваны обучением.
Оба исследования пластичности выполнялись на млекопитающих, и поэтому они с большей вероятностью имеют отношение к нервной системе человека, чем исследования, проводимые на лягушках и других земноводных. С земноводными экспериментальные исследования проводились в широких масштабах. Наиболее яркие проявления нейронной пластичности (включая модификацию отображения посредством эффекта типа "резиновой пленки") изучались для земноводных еще задолго до того, как аналогичный эффект был обнаружен Уоллом и Эггером у взрослых крыс. Результаты исследований на земноводных достаточно полно отражены в книге Гейза [8].
Пластичность в нервной системе земноводных
Нервную систему земноводных можно исследовать такими экспериментальными методами, которые неприемлемы в случае млекопитающих. Нервная система животных состоит из двух частей - центральной нервной системы, которая включает головной мозг, спинной мозг, зрительные нервы и сетчатку, и периферической нервной системы, состоящей из отростков центральной нервной системы, которые соединяются с мышцами и органами чувств. У млекопитающих повреждение центральной нервной системы не ликвидируется структурно: благодаря какой-либо форме пластичности возможно некоторое восстановление функции, то перерезанный зрительный нерв млекопитающего никогда не восстанавливается. В этом отношении нервная система земноводных ведет себя совершенно иначе: соединения в центральной нервной системе, поврежденные в результате хирургического вмешательства, могут восстанавливаться.
Сетчатка каждого глаза лягушки отображается на зрительную долю противоположной стороны мозга. Если зрительный нерв перерезан, но концы находятся довольно близко друг от друга, то со временем они соединяются - и зрение в пораженном глазу лягушки восстанавливается. В этом мы можем убедиться, замечая, что лягушка в состоянии поймать язычком приманку, которой перед ней помахивают, держа в различных частях зрительного поля. Проверка с помощью микроэлектрода также показала, что произошло восстановление отображения в зрительной доле. Восстановление наблюдается даже в том случае, если концы зрительного нерва "разлохматить". Мысль о том, что волокна "находят дорогу" к своим прежним соединениям на другой стороне среза, интуитивно кажется весьма нелепой: ведь для этого волокна должны содержать какие-то химические вещества, обеспечивающие их правильное соединение. Многим исследователям более разумным представляется другое объяснение: волокна соединяются случайным образом, а восстановление зрения происходит в результате определенного процесса обучения на зрительном опыте.
Однако некоторые дальнейшие эксперименты, проведенные Сперри [9], заставили отказаться от этой гипотезы. Было установлено, что если во время перерезания зрительного нерва глаз был повернут на 180°, то зрение восстанавливается, но изображение оказывается повернутым на 180° (и остается таким в дальнейшем). В том, что зрительное восприятие становится "перевернутым", можно убедиться, если помахивать приманкой в различных частях зрительного поля; мы увидим, что лягушка стреляет язычком в неверном направлении. К подобному заключению приводит и исследование микроэлектродными методами отображения на зрительной доле.
Можно также прервать оба зрительных нерва и дать им соединиться так, чтобы нерв, который шел к правому глазу, теперь соединился с левым, и наоборот. И снова зрение восстанавливается, но внутренняя (назальная) сторона правого глаза отображается в то место на правой зрительной доле, которая прежде представляла внутреннюю (назальную) сторону левого глаза, и т. д. Зрение оказывается обращенным слева направо (рис. 41).
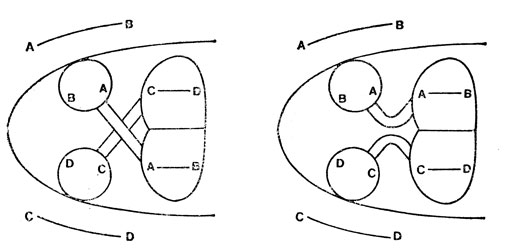
Рис. 41. Результат устранения перекрещивания двух зрительных нервов (нервы перерезаются, при этом создается препятствие восстановлению прежних соединений). Глаза изображены кружками. Точки А, В, С и D зрительного поля образуют изображения на соответствующих сетчатках. После операции точки А и В отображаются на тектум. там, где раньше отображались точки С и D. Приманка, показанная в точке А, заставляет лягушку ударить в точку С, а приманка, помещенная в точку В, заставляет лягушку ударить в точку D. Это говорит о повороте изображения на 180° слева направо
Эти результаты, казалось, отвечали точке зрения Сперри, утверждающего, что нервные волокна способны "распознавать" свои прежние соединения (или в случае перекрещивания зрительных нервов распознавать в другом нерве волокна, соответствующие прежним соединениям) вследствие какой-то их химической специфичности. Создается впечатление, что в целом все предопределено генетически, и поэтому нет необходимости постулировать существование какой-то формы пластичности.
Но, как показали дальнейшие эксперименты, картина не столь проста. В них было установлено, что в результате хирургического вмешательства какая-то часть сетчатки может оказаться отображенной на всю зрительную долю или, наоборот, часть зрительной доли может содержать отображение всей сетчатки. Эффект "резиновой пленки" может работать с той и другой стороны. (Во многих отчетах по этой работе для обозначения зрительных долей использовалось слово тектум.)
Гейз разработал полезную методику, связанную с формированием составных глаз (которые, однако, не следует путать с многоячеистыми составными глазами насекомых). Гейз имел в виду глаз, "образованный" из двух половинок, взятых от левого и правого глаз. В результате операции он создавал у животных двойной назальный или двойной височный глаз. Двойной назальный глаз состоит из назальной (внутренней) половины правого глаза и назальной половины левого глаза. Подобным же образом двойной височный глаз состоит из двух височных (внешних) половин правого и левого глаз. Оказывается, вполне реально трансплантировать глаза (или половины глаз) одного животного другому, получая указанные результаты.
В одной серии экспериментов Гейз формировал составные глаза у земноводных на ранней стадии их развития - до того, как образовывались соединения, идущие от глаз к тектуму. Когда затем эти соединения возникали, обнаруживалось, что каждый "полуглаз" отображен на всю зрительную долю противоположной стороны, как показано на рис. 42. Каждая точка зрительной доли (за исключением складки в точке В) соответствует двум точкам на сетчатке. Ясно, что принцип химической специфичности действует не совсем так, как предполагал Сперри.
![Рис. 42. Отображение зрительного поля - сначала на сетчатке, а затем на тектуме - у лягушки с нормальным глазом (N), с двойным назальным глазом (NN) и с двойным височным глазом (ТТ). (По данным Гейза [8])](pic/000051.jpg)
Рис. 42. Отображение зрительного поля - сначала на сетчатке, а затем на тектуме - у лягушки с нормальным глазом (N), с двойным назальным глазом (NN) и с двойным височным глазом (ТТ). (По данным Гейза [8])
Модель "торговцы чаем"
Уилшоу и Ван-дер-Малсбург [10] предложили теорию, объясняющую описанные здесь экспериментальные результаты (за исключением колатеральной проекции, о которой речь пойдет ниже). Согласно этой теории, волокна зрительного нерва не имеют особой "пометки", которая указывала бы на наличие в каждом волокне некоего специфического химического вещества. Вместо этого в отдельных точках сетчатки образуется сравнительно небольшое число различных веществ, которые, диффундируя из мест своего образования, распределяются по сетчатке таким образом, что их концентрация уменьшается пропорционально расстоянию. Каждое конкретное волокно несет в себе смесь "маркерных веществ", соответствующую относительной концентрации этих веществ в том месте сетчатки, откуда берет начало данное волокно.
Такие химические смеси, переносимые нервными волокнами, дают им возможность формировать непрерывное отображение на зрительной доле, как только они впервые достигают ее в ходе эмбрионального развития животного. В своей более ранней работе [11] Уилшоу и Ван-дер-Малсбург попытались объяснить, как формируется такое отображение, предложив для этой цели модель "торговцы чаем" (см. ниже). Волокна нерва, перенося химические смеси на зрительную долю, формируют на ней распределение веществ, подобное распределению этих веществ на сетчатке. Если зрительный нерв перерезается, но ему предоставлена возможность регенерировать, то отображение восстанавливается в силу того, что каждое волокно соединяется со зрительной долей в нескольких точках, но соединение "крепче" там, где химический состав самого волокна в большей степени соответствует химическому составу данной области зрительной доли. Соединения, плохо согласованные, исчезают, а рядом с уже имеющимися соединениями формируются новые. Это позволяет "главному соединению" данного волокна перемещаться к той точке на зрительной доле, которая химически наиболее сходна с нервным волокном. Распределение маркерных веществ на зрительной доле составляет память, обеспечивающую упорядоченное восстановление соединений.
Установлено, что если ввести некоторый верхний предел для суммы синаптических весов каждого волокна, то эта модель объясняет и все наблюдаемые эффекты "резиновой пленки". Обратимся теперь к аналогии с торговцами чаем, которые приезжают из Индии в Англию, чтобы организовать торговлю чаем. В этой модели территория Индии представляет собой сетчатку, а территория Англии - поверхность зрительной доли. Предполагается, что в Индии чай выращивают на каком-то числе плантаций, причем чай с каждой плантации отличается своим вкусом. Торговцы приезжают из разных деревень, где составляются свои смеси чая. Состав чайной смеси, характерный для какой-то конкретной деревни, содержит исходные виды чая в таких пропорциях, которые обратно пропорциональным образом связаны с расстоянием от деревни до соответствующей плантации.
Торговцы в состоянии привести с собой лишь какое-то определенное суммарное количество чая. У покупателей вырабатывается предпочтение той смеси чая, которую они обычно покупают, и торговец большую часть своего чая поставляет тем покупателям, которым этот чай по душе. Таким образом, покупатель закрепляет за собой право на приобретение нужного ему сорта чая. Но вместе с тем каждый торговец, конкурируя со своими коллегами, выделяет и распространяет определенное количество чая "на пробу", с тем чтобы привлечь новых покупателей. Моделирование на вычислительной машине показало, что такое сочетание взаимодействия и конкуренции может создать некоторое гладкое отображение при условии, что в начальных контактах торговцев с покупателями имеется хотя бы небольшое соответствие в порядке обслуживания.
В случае зрительного нерва соответствие в порядке обслуживания обеспечивается благодаря тому, что по мере роста из сетчатки в пучке волокон сохраняется определенный порядок. Сначала формируется отображение между относительно малыми областями сетчатки и зрительной доли, но оно сохраняется и в дальнейшем, по мере увеличения числа волокон, идущих от сетчатки к зрительной доле. В модели это соответствовало бы следующей ситуации. Потребление чая начинается с небольшой области Англии, куда он доставляется также из какого-то небольшого района Индии, а затем оно постепенно распространяется по всей стране. Это в свою очередь способствует тому, что выращиванием чая начинают заниматься и в других районах Индии.
Модель "торговцы чаем" и исследования по имитации ее на вычислительной, машине обычно не связывают с искусственным интеллектом, поскольку эти исследования не столь целенаправленны, как большинство работ в области искусственного интеллекта. Тем не менее эти исследования дают общее представление о некотором классе проявлений нейронной пластичности и, безусловно, вносят значительный вклад в понимание процессов обработки информации, происходящих в мозге.
Колатеральная проекция
Основная проекция сетчатки на зрительную долю носит контралатеральный характер: сетчатка каждого глаза соединяется со зрительной долей противоположной стороны головы. У лягушки (в отличие от человека) наблюдается полное перекрещивание - все волокна от правого глаза идут в левую долю и наоборот.
Но несмотря на полное перекрещивание, лягушка способна связывать между собой изображения одной и той же точки, сформированные на двух сетчатках. Это важно, если лягушка обладает бинокулярным зрением (обеспечивающим стереоскопическое восприятие глубины). Соответствующие точки на двух зрительных долях действительно связаны между собой, причем в обеих направлениях. В результате возбуждение какой-то точки на любой из сетчаток создает реакцию в зрительной доле, расположенной с той же стороны (колатеральная доля); при этом сигнал сначала идет к контралатеральной доле, а затем по пути, соединяющему две доли.
В противоположность первичной контралатеральной проекции соединительные пути колатерального типа, оказывается, реагируют таким образом, что это можно объяснить, лишь предположив, что они формируются в процессе накопления животным зрительного опыта. Когда один глаз животного поворачивают так, чтобы проекция сетчатки на зрительную долю создавала перевернутое изображение окружающего мира, то обнаруживается, что волокна, соединяющие две доли, связывают точки, соответствующие одной и той же точке в реально наблюдаемом зрительном поле. Способность к обучению, которая, как ни странно, отсутствует в контралатеральной проекции, проявляется в колатеральной проекции. Таким образом, в смежных частях нервной системы лягушки действуют два совершенно разных типа нейронной пластичности.
Нейрофизиология и искусственный интеллект
Мы довольно подробно рассмотрели вопросы нейрофизиологии зрения и нейронной пластичности. Экспериментальные результаты, полученные при исследовании зрительной системы животных, в принципе имеют отношение к системам искусственного интеллекта: по крайней мере нетрудно заметить удивительное соответствие между рис. 16 и 17, относящимися к работе Паркса по распознаванию символов, и рис. 40.
Исследования по нейронной пластичности скорее могут быть связаны с "кибернетическим" подходом к искусственному интеллекту, т. е. подходом с позиций "самоорганизующихся систем". Основное же направление работ в области искусственного интеллекта, очевидно, следует связывать с функциями мозга на более высоком, организационном, уровне. Однако оба этих уровня должны в конце концов сомкнуться. Как отмечалось в гл. 1, "экспериментальная эпистемология" Маккалока была направлена именно на то, чтобы добиться гармонического сочетания этих двух подходов.
Как мы могли заметить, идеи пластичности в настоящее время активно развиваются, и в этой области, безусловно, наблюдается определенный прогресс. Возможно, что недалек тот час, когда будет сделан существенный шаг в направлении объединения двух подходов к исследованию искусственного интеллекта, к чему и призывал Маккалок.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'