2.2. Мозг как нейронная сеть
* (В этом разделе разъясняются такие основные понятия нейрофизиологии, как аксон, дендрит, синапс и пороговое значение, и читатель, хоть немного знакомый с нейрофизиологией, может его не читать.)
В разд. 1.2 мы уже познакомились с основными "компонентами" рецепторов, нейронов и эффекторов, схематически представленных на рис. 3. В мозге многие миллиарды нейронов связаны между собой, образуя цепи, петли и сложно переплетенные сети, так что сигналы, поступающие в сеть от рецепторов, взаимодействуют там с миллиардами других сигналов, полученных системой раньше, и в результате этого взаимодействия изменяется состояние системы и возникают сигналы, возбуждающие эффекторы. Благодаря этому поведение организма в данный момент определяется не только поступившими в этот момент стимулами, но и всем прошлым опытом, выражающимся в состоянии (и изменении структуры) его нейронной сети (рис. 4).
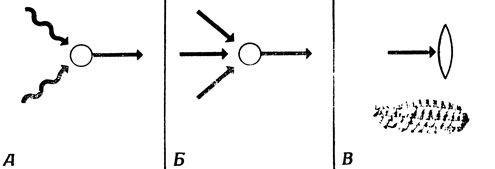
Рис. 3. Основные элементы, которые необходимо знать при изучении нервной системы. А. Рецепторы - преобразуют энергию в 'невральные сообщения'. Б. Нейроны - преобразуют полученные сообщения в новые сигналы. В. Эффекторы - например, мышечные волокна, сокращающиеся под действием полученных сигналов
Все эти соображения можно суммировать, сказав, что в нашей теории мы рассматриваем мозг как нейронную сеть; тем самым в известном смысле устанавливается некий критерий для оценки объяснений, из которых слагается наша теория мозга. Другими словами, в нашей теории мозга (но, конечно, не в нашем исследовании искусственного интеллекта) основной упор делается, как уже отмечалось в рассуждениях о различных уровнях описания в разделе 1.2, на изучение свойств систем взаимосвязанных нейронов, а не на чисто психологические вопросы и не на исследование биохимических систем. Мы сосредоточим свое внимание на некоторых свойствах нейронов и даже более крупных структур головного мозга и будем строить теорию мозга на этой основе, а не на основе, например, биохимического изучения воздействия разных лекарственных веществ на функции нервной системы. Мы не будем также уделять внимание другим клеткам мозга, например глиальным клеткам, которые играют важную роль в метаболизме, а, по мнению некоторых исследователей, возможно, принимают участие и в формировании памяти.
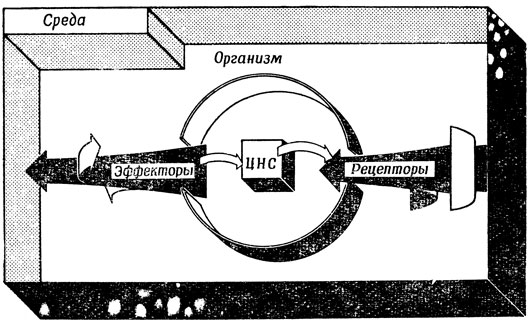
Рис. 4. Взаимодействие между организмом и средой. Реакция рецепторов на воздействия среды и сигналы обратной связи от эффекторов преобразует поведение ЦНС (центральной нервной системы), которая, в частности, управляет эффекторами, обеспечивающими активность организма
Несмотря на такую ограниченность нашего подхода, мы надеемся решить с его помощью много важных задач. И при этом вовсе не исключаем возможности слияния в будущем нашего подхода с другими подходами, например основанными на изучении химической природы процессов мышления.
Всем нам известны пять основных чувств: слух, зрение, осязание, вкус и обоняние. Специализированные рецепторы глаза реагируют на свет; рецепторы, расположенные во внутреннем ухе, через посредство ряда структур воспринимают колебания давления воздуха; некоторые рецепторы кожи реагируют на изменение давления, а другие, расположенные на языке и в полости рта, - на самые малые концентрации химических веществ. Но, несмотря на всеобщее убеждение, что шестое чувство дано далеко не каждому, у всех нас есть и шестое, и седьмое чувство и еще много других. Помимо "классических" осязательных рецепторов в коже имеются и другие, реагирующие на температуру или на болевые воздействия, например на нарушение целостности ткани.
Кроме внешних рецепторов имеются и рецепторы в мышечной ткани, которые регулируют работу мышц, сухожилий и суставов и постоянно обеспечивают сигналы "обратной связи", несущие информацию о напряжении, развиваемом мышцей, ее длине, суставном угле и т. п., поскольку для нормального функционирования эффекторов необходимо знать не только то, к чему должна стремиться система, но и то, где она сейчас находится. Вестибулярный аппарат, расположенный в височной кости, контролирует положение тела в пространстве и ускорение его движения. Кроме того, во внутренних органах имеются рецепторы, контролирующие химический состав крови, кровообращение, пищеварение и другие функции. В мозге имеются также рецепторы, сигнализирующие о недостаточном содержании в крови воды, сахара и т. п. При раздражении соответствующих клеток мозга животное без конца пьет воду или поглощает огромные количества пищи, хотя у него нет объективных оснований для жажды или голода. Это происходит потому, что мозг получил сигнал о недостатке воды или пищи и дал животному соответствующие инструкции, несмотря ни на какие противоречащие этому сигналы, которые могут поступать из переполненного желудка.
Для того чтобы лучше понять, что происходит на пути от рецептора к эффектору, нужно ближе познакомиться со свойствами нейрона. Но, оказывается, в природе вообще не существует того, что можно было бы назвать типичным нейроном (рис. 5). Поэтому нам остается только схематически описать свойства, общие для большинства нейронов, отметив, что в каждом специальном случае эту обобщенную картину придется соответственно модифицировать.
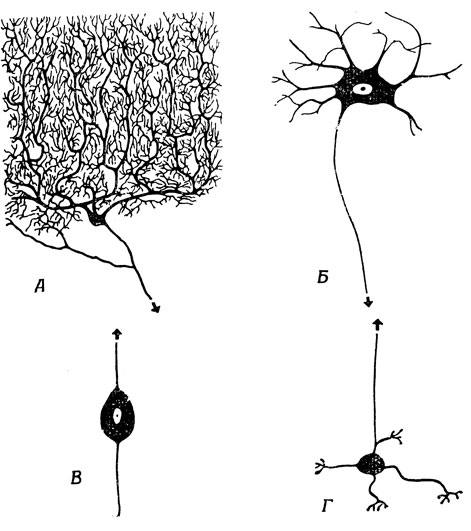
Рис. 5. Этот рисунок напомнит нам, сколь схематично идеализированное представление о нейроне (рис. 6). Приведены всего 4 из тысяч различных типов нейронов. Во всех случаях стрелы направлены к концевым разветвлениям аксонов, не показанных на рисунках. А. Клетка Пуркинье мозжечка. Б. Двигательный нейрон (наиболее близкий к идеальному нейрону). В. Биполярный нейрон обонятельной системы. Г. Клетка-зерно мозжечка
Основные черты нейрона можно видеть на рис. 6, где схематически изображен мотонейрон спинного мозга млекопитающего. От тела нейрона (или сомы) отходит несколько ветвящихся отростков, называемых дендритами (от греч. дендрон, что означает "дерево"). Эти дендриты образуют основную часть входной поверхности нейрона. От тела клетки в точке, называемой аксонным холмиком (переходящим в начальный сегмент), отходит также длинное волокно, называемое аксоном, который образует обычно так называемые концевые разветвления. Концевые утолщения ветвей аксона, или пуговки, располагаются на других нейронах или на эффекторах. Места соприкосновения этих веточек с клеткой, на которой они лежат, называются синапсами.
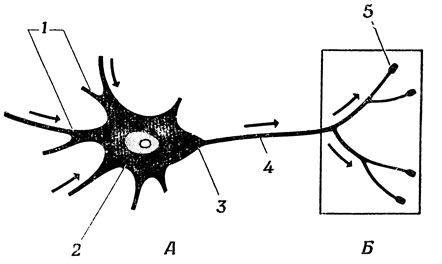
Рис. 6. Схема нейрона. Возбуждение рецепторов и других нейронов изменяет мембранный потенциал дендритоз (1) и тела клетки (2). Эффекты этих изменений сходятся на аксонном холмике (3), в результате чего - при соответствующей пространственно-временной конфигурации пришедших сигналов - импульс мембранного потенциала начинает распространяться по аксону (4), расходится но концевым разветвлениям и активизирует синаптические концевые луковички (5), которые в свою очередь изменяют мембранный потенциал других нейронов или мышечных волокон
Удобнее всего представить, что поток информации от нейрона к нейрону происходит в направлении, указанном на рис. 6 стрелками; хотя аксон и может "проводить" в обоих направлениях, в большинстве случаев возбуждение передается на дендрит или сому той клетки, с которой он образует синапс. При этом возбуждение передается сначала на холмик, а затем распространяется по аксону, пока не достигнет концевого разветвления. Некоторые аксоны очень длинные. Например, нейрон, управляющий движением большого пальца ноги, находится в спинном мозге и, следовательно, длина его аксона не меньше длины ноги человека. Длина аксонов подобных нейронов просто огромна по сравнению с размерами нейронов головного мозга, которые настолько малы, что в относительно^ скромном объеме черепа их умещается более десяти миллиардов. Например, в зрительной системе имеются крошечные клетки, называемые амакриновыми, ответвления которых нельзя разделить на дендриты и аксоны, так как они очень короткие и могут проводить возбуждение в обе стороны, выполняя роль локальных модуляторов для соседних нейронов.
Для того чтобы лучше разобраться в этой системе "связи", удобно рассматривать клетку как живое существо, заключенное в мембрану, между наружной и внутренней поверхностями которой существует разность потенциалов. Если изменить эту разность потенциалов между внешней и внутренней поверхностями мембраны, то это изменение распространяется пассивным образом, примерно так же, как происходит распространение тепла по металлическому стержню. Изменение температуры на одном конце стержня может передаваться и другим его участкам, но с увеличением расстояния от точки нагрева изменение температуры становится все менее выраженным (рис. 7). Точно так же обычное изменение мембранного потенциала пассивно распространяется по волокну, и чем больше мы удаляемся от точки исходного возбуждения, тем позже оно наступает и тем слабее выражено.
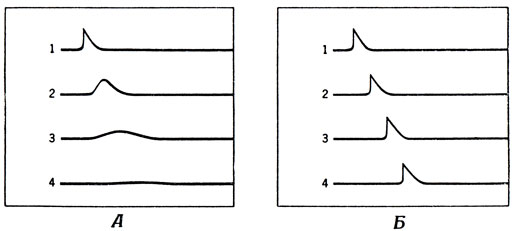
Рис. 7. Два основных типа распространения возбуждения по нейрону. А. Пассивное распространение изменений мембранного потенциала (аналогично распространению тепла по металлическому стержню) - пик сигнала постепенно затухает; такая проводимость характерна для дендритов, тела клетки и очень коротких аксонов. Б. Активное распространение - возбуждение в одном участке вызывает возбуждение в следующему характерно для передачи сигналов по длинным аксонам; было обнаружено также и в дендритах некоторых клеток, например клеток Пуркинье. Цифрами обозначены последовательные моменты времени
Для клеток с короткими отростками (типа палочек, колбочек и вставочных нейронов сетчатки) этого пассивного распространения достаточно для того, чтобы передавать сигналы о6 изменении потенциала из одного конца системы в другой. Но если аксон достаточно длинный, то этого механизма совершенно недостаточно, поскольку возбуждение на одном конце затухнет полностью, так и не добравшись до другого.
К счастью, у клеточных мембран есть дополнительное свойство, состоящее в том, что если изменение разности потенциалов оказывается достаточно большим (или, как говорят, превышает порог), то в цилиндрических образованиях типа аксона возникает импульс, который активно распространяется, сохраняя постоянную амплитуду, т. е. не затухает по мере распространения. Для того чтобы понять, как это происходит, представьте себе металлическую трубку, начиненную порохом. Если такую трубку слегка нагреть, то порох не воспламеняется, и разность температур распространяется пассивно, постепенно затухая. Если, однако, превысить температуру возгорания пороха, то в нагреваемом участке трубки порох вспыхнет и выделяемого в результате этого тепла оказывается достаточно, чтобы поджечь порох на соседнем участке и так далее по всей длине трубки. В этом случае мы имеем дело уже не с пассивным распространением тепла, полученного при нагревании. Сообщенное тепло послужило лишь толчком к возникновению "регенеративного" процесса - цепной реакции, которая сама обеспечивает себя энергией, и стоит нам вызвать эту реакцию в одном месте, как она высвобождает энергию, запасенную в соседнем участке, которая в свою очередь вызывает реакцию на следующем участке, и так далее. Аналогичным образом обстоит дело и с мембраной нервного волокна. Так, если разности потенциалов на дендритах и теле нейрона создают (обычно в результате пассивного распространения возбуждения на мембране аксонного бугорка) разность потенциалов, превышающую определенный порог, то запускается регенеративный процесс, т. е. возбуждение в одном участке оказывается достаточным, чтобы запустить аналогичный процесс в соседнем участке, так что возникает незатухающий импульс, распространяющийся вдоль аксона.
В нашем примере с запальным устройством мы можем долбиться активного распространения лишь однажды: после того как весь порох выгорит, останется лишь металлическая трубка, способная проводить тепло только пассивно. Поэтому аксон лучше сравнивать с перезаряжаемым запалом: представим себе, что после распространения каждого такого импульса по аксону происходят химические процессы, эквивалентные перезарядке запала. (Эта функциональная эквивалентность не означает, что изменения, происходящие в мембране, действительно аналогичны перезарядке!) Поэтому имеется короткий рефрактерный период, во время которого аксон не может передавать новый им - импульс; именно в этот период и происходит указанное химическое восстановление.
Если бы мы нанесли раздражение в каком-либо участке по ходу аксона, то возбуждение распространялось бы в обоих направлениях. Однако если импульс возникает на одном конце аксона (обычно на аксонном холмике), то он может распространяться лишь в одном направлении, потому что возбужденный участок аксона становится рефрактерным и остается в этом состоянии еще достаточно долго. Перемещаясь по аксону, импульс вызывает возникновение новых импульсов в каждом из ветвлений (или коллатералей) аксона, а те в свою очередь вызывают новые импульсы в каждой из еще более тонких веточек (рис. 8). Когда же импульс доходит до концевой пуговки, то после небольшой задержки на мембране клетки, с которой аксон образует синапс, возникает разность потенциалов. Мембрана концевой пуговки называется пресинаптической мембраной, а мембрана той клетки, на которую передается импульс, - постсинаптической мембраной.
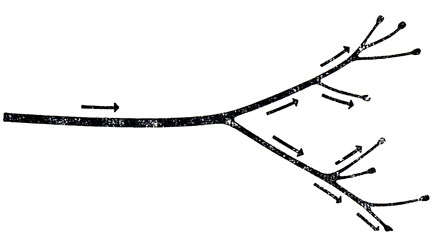
Рис. 8. Обычно возбуждение распространяется от аксэнного холмика по направлению к разветвлению: дойдя до точки ветвления спайк возбуждает спайк во всех отходящих от этой точки ветвях
Как это ни удивительно, в большинстве случаев непосредственная причина изменения потенциала постсинаптической мембраны имеет не электрическую, а химическую природу. В некоторых особых синапсах, которые отличаются большими размерами, или в таких синапсах, для которых характерно тесное соприкосновение мембран, изменение поляризации постсинаптической мембраны происходит без участия химических медиаторов. Однако, как правило, электрический импульс, достигнув, концевой пуговки вызывает выделение из так называемых синаптических пузырьков вещества, называемого медиатором, которое диффундирует затем через небольшую синаптическую щель (рис. 9) к постсинаптической мембране. Попав на эту мембрану, медиатор вызывает изменение ее поляризации. Медиатор по своему действию может быть двух основных типов:
возбуждающим - если он изменяет разность потенциалов на постсинаптической мембране в направлении к пороговому значению, или тормозным, т. е. изменяющим разность потенциалов в обратную сторону.
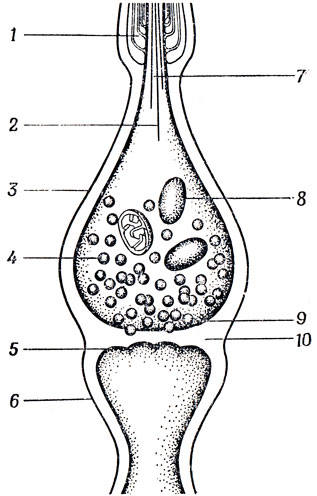
Рис. 9. Схема 'типичного' синапса. Пришедший по аксону (7) импульс не оказывает прямого электрического воздействия 2 на нейрон, с которым этот аксон контактирует, а вызывает освобождение некоторого числа синаптических пузырьков (4), в которых заключен медиатор; эти пузырьки диффундируют через пресинаптическую мембрану (9) и синаптическую щель (10) и деполяризуют (или гиперполяризуют) постсинаптическую мембрану (5). В данном случае постсинаптическая мембрана образует поверхность крошечного шипика дендрита, обеспечивающего синаптический контакт. Нейрофиламенты (2) и митохондрии (8) принимают участие в метаболизме нервной системы. Миелиновая оболочка (1), которой одето большинство аксонов, служит, помимо всего прочего, для увеличения с скорости распространения спайков
Единственное обобщающее высказывание, которое можно сделать относительно нервной системы, звучит примерно так: "О нервной системе нельзя делать общих утверждений, за исключением того, что о нервной системе нельзя делать общих утверждений, за исключением того, что... и т. д.". Однако многие нейрофизиологи принимают закон Дейла, согласно которому если проследить за всеми ветвлениями аксона какого-либо одного нейрона млекопитающего, то все синапсы, образуемые этим аксоном с другими клетками, окажутся либо только возбуждающими, либо только тормозными. К беспозвоночным это утверждение неприменимо. Например, Кандель и др. [133] установили, что у моллюска Aplysia можно выделить вставочный нейрон, различные ветвления которого осуществляют противоположные синаптические воздействия. "Закон" Дейла может оказаться неверным даже для млекопитающих, поскольку имеются косвенные данные [224], что в спинном мозге кошки примерно 30% волокон образуют и возбудительные и тормозные синапсы (см, также [225]). Естественно, что на любой данной клетке могут быть синапсы как того, так и другого типа, поскольку они образованы многими разными нейронами.
Многочисленные незначительные изменения мембранного потенциала отдельной клетки, вызванные активностью всех ее синапсов, пассивно (в большинстве случаев, но далеко не всегда) распространяются по мембране дендритов и тела клетки, постепенно затухая. Но в аксонном холмике эти потенциалы сходятся, и хотя каждый из них может быть очень слабым, суммарное возбуждение может во много раз превышать порог (рис. 10).
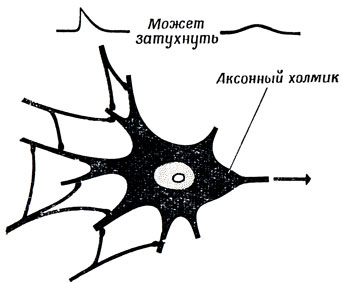
Рис. 10. Эффект каждого отдельного синаптического возбуждения может быть слабым, однако совокупный эффект, получающийся в результате пространственной и временной суммации сотен таких отдельных эффектов, может оказаться выше порогового и возбудить импульс в аксонном холмике
Другими словами, импульс, достигающий концевой пуговки аксона, обычно вызывает лишь подпороговое возбуждение постсинаптической мембраны. Тем не менее согласованный эффект многих таких подпороговых изменений в аксонном холмике может превысить порог, и если это происходит в момент, когда рефрактерный период после предыдущего возбуждения уже закончился, то вдоль аксона начнет распространяться следующий импульс.
Для простоты представим себе схематически эффект синаптического возбуждения как изменение во времени потенциала в направлении порогового значения по типу первого импульса, показанного на рис. 11. (Мы не будем здесь изображать тормозный эффект, считая, что он просто препятствует изменению потенциала в направлении к порогу, либо внося в суммарный эффект "отрицательный вклад", либо "шунтируя" возбуждение.) Изменения разности потенциалов, вызванные одним спайком, пришедшим к синапсу, почти всегда будут гораздо ниже порогового. Но здесь возможен кумулятивный эффект, эти изменения могут суммироваться с изменениями, вызванными предшествующим спайком, которые еще не успели затухнуть. Поэтому, если подпороговые изменения поступают одно за другим с достаточно короткими интервалами, они могут накапливаться и в конечном счете превысить порог (рис. 11), за другим. Если на клетке имеется порядка 10 000 синапсов, то можно ожидать, что на протяжении одного рефрактерного периода возбудится по крайней мере тысяча из них. В связи с этим возникают сложные взаимодействия различных событий, происходящих на протяжении как одного, так и разных рефрактерных периодов. Например, из рис. 11 видно, что вспышки активности нейрона в моменты времени t3 и t6 одинаковы, однако спайк генерируется лишь в момент t6, хотя для этого потенциал должен был значительно превысить нормальный порог, так как он пришелся на относительный рефрактерный период, последовавший за спайком, генерированным в момент t4.
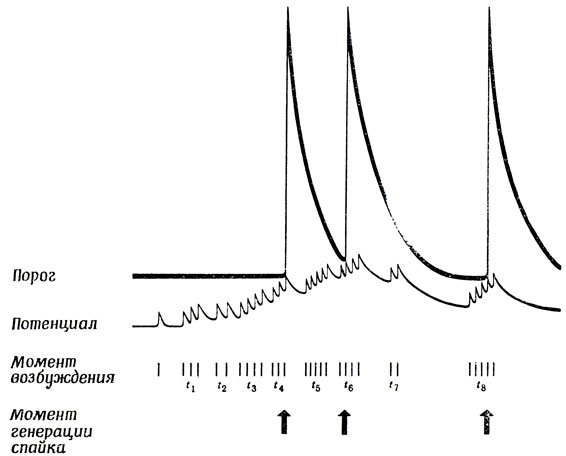
Рис. 11. Схема временной суммации. Каждое возбуждение вносит небольшой вклад в мембранный потенциал, который, затухая, быстро возвращается к своему уровню покоя. Однако, если интервалы между возбуждениями достаточно малы, их суммы может оказаться достаточно для того, чтобы перекрыть порог. Резкое повышение порога, следующее за генерацией каждого спайка, позволяет понять, почему аксон не способен проводить два импульса, если интервал между ними меньше рефрактерного периода
Масштабы синаптического взаимодействия клеток лучше всего иллюстрировать на примере клеток Пуркинье мозжечка (рис. 12) с весьма многочисленными и сильно ветвящимися дендритами. Клетки многих типов образуют синапсы с клеткой Пуркинье, и эти синапсы весьма разнообразны. Например, от других клеток мозжечка, так называемых клеток-зерен, отходит примерно 200 000 волокон, идущих практически параллельно друг другу. Прорезая массу дендритов клетки Пуркинье, они образуют с ними синапсы, по одному на аксон каждой клетки- зерна. Другие волокна, которые называют лазящими, в самом деле как бы лазают по наиболее густым участкам дендритного ветвления клетки Пуркинье, образуя с ней многочисленные синапсы. Поэтому, если лазящее волокно командует клетке Пуркинье возбудиться, та послушно возбуждается. Согласованное действие многих параллельных волокон вкупе с другими волокнами может регулировать уровень этого возбуждения или возбуждать клетку Пуркинье непосредственно. Все это еще раз напоминает нам о разнообразии синапсов по типу, размеру и форме.
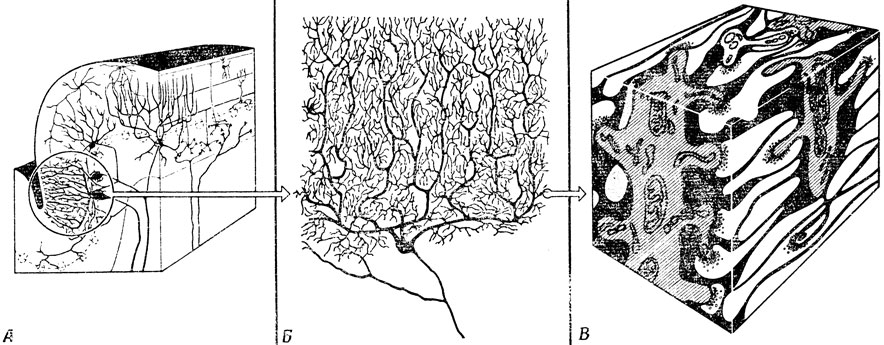
Рис. 12. Три схемы, дающие некоторое представление о сложности организации нервной системы. Мозжечок состоит из нескольких долей, каждая из которых в свою очередь делится на несколько листков. А. Участок одного листка; изображена лишь небольшая часть составляющих его клеток. Б. Более подробная схема одной из таких клеток - клетки Пуркинье; ее площадь в плоскости среза составляет примерно 0,3 мм2, а толщина - не более 0,03 мм. Она образует синапсы с одним лазящим волокном, с 200 000 параллельных волокон и с аксонами многих других клеток мозжечка. В. Показана часть одной из самых мелких дендритных веточек клетки Пуркинье. Можно видеть, что от этой веточки отходят семь синаптических шипиков (пять из них приходятся на переднюю грань), вступающих в контакт с проходящими вблизи параллельными волокнами
Геометрия различных синапсов, образуемых данной клеткой, определяет, сложатся ли временные соотношения разных синаптических активностей таким образом, чтобы суммарный эффект затухающих изменений потенциала, поступающих на аксонный холмик, превысил пороговое значение; поэтому в зависимости от геометрии синапсов, взаимодействие между клетками может быть самым различным. В разд. 2.4, рассмотрев роль нейронов в предварительной обработке зрительной информации, мы начнем лучше разбираться во всех тонкостях их функционирования. Здесь же мы попытаемся связать то, что мы рассказали о нейронах, со свойствами рецепторов и эффекторов.
Что касается организации "входа" нервной системы, то рецепторы, так же как и нейроны, обладают свойством генерировать потенциалы, передающиеся различным синапсам на поверхности нейронов. Однако "входная" поверхность рецепторов не образует синапсов с другими нейронами; она может лишь преобразовывать энергию, поступающую из внешней среды, в изменение мембранного потенциала, которое в свою очередь может распространяться пассивно или активно (многие рецепторы, например зрительные, не генерируют спайков). Палочки и колбочки глаза содержат различные пигменты, способные химически реагировать на свет, а эта химическая реакция в свою очередь вызывает локальные изменения мембранного потенциала, который называют генераторным. При достаточной интенсивности света, падающего на колбочку или палочку, эти изменения потенциала вызовут изменения потенциала вставочных нейронов, которые могут оказаться достаточно сильными для того, чтобы возбудить ганглиозные клетки сетчатки (см. разд. 2.4): таким образом сообщение о свойствах света будет передано в нервную систему в виде серии импульсов. Другие рецепторы могут преобразовывать в генераторные потенциалы давление и т. п.
Для того чтобы перейти к обсуждению организации "выхода" нервной системы, мы должны прежде всего вспомнить о том, что мышца представляет собой совокупность многих тысяч мышечных волокон. Нейроны, управляющие мышечными волокнами, называют двигательными нейронами, или мотонейронами; они лежат в спинном мозге, и их аксонам иногда приходится проходить огромные (по нейронным стандартам) расстояния, прежде чем они могут образовать синапс с мышечным волокном. Таким образом, наименьшая функциональная единица на выходе нервного тракта (по крайней мере для млекопитающих) - это двигательная единица, состоящая из тела мотонейрона, его аксона и группы мышечных волокон, управляемых этим аксоном.
Мышечное волокно сходно с нейроном в том отношении, что и оно получает входные сигналы от других клеток, с которыми оно образует синапсы. Однако в ответ на такую активизацию оно не генерирует импульсы, которые бы распространялись по аксону, а просто сокращается; поэтому мотонейроны, образующие синапсы на мышечном волокне, по структуре своей импульсации определяют степень сокращения мышцы, образованной из этих волокон, и в конечном счете управляют движением. (Аналогичные замечания можно сделать и о клетках, которые выделяют различные химические вещества в кровь или пищеварительный тракт или же секретируют пот или слезную жидкость.)
Так как нейрон не может послать мышце команду "расслабиться" (максимум, что нейрон может сделать, - это перестать посылать мышце команду сокращаться), мышцы животных, как правило, сгруппированы в антагонистические пары (рис. 13). Сокращение одной из мышц такой пары вызывает расслабление другой. Так, одна группа мышц заставляет нас разогнуть руку в локте, а другая - согнуть ее. И если нам нужно разогнуть руку, мы не командуем мышцам-сгибателям расслабиться, просто перестаем заставлять их сокращаться, после чего они автоматически расслабляются при сокращении мышц-разгибателей.
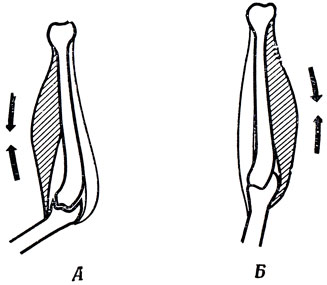
Рис. 13. Схематическое изображение локтевого сустава, показывающее антагонистическое действие мышц-сгибателей и мышц-разгибателей. А. Сокращение мышцы-сгибателя уменьшает суставной угол. Б. Сокращение мышцы-разгибателя увеличивает суставной угол
Раздражение двигательной концевой пластинки (т. е. синапса мотонейрона на мышечном волокне) вызывает кратковременное одиночное сокращение мышечного волокна. Если импульсы приходят на концевую пластинку с низкой частотой, то это вызывает серию таких сокращений, при каждом из которых механическая реакция продолжается дольше, чем стимул, т. е. потенциал действия. С повышением частоты раздражения наступает момент, когда второй импульс поступает раньше, чем закончится механическая реакция на первый. А это вызывает механическую суммацию одиночных сокращений. До известного предела эффект суммации возрастает по мере уменьшения интервала между импульсами. Однако, как только эти интервалы приблизятся по продолжительности к рефрактерному периоду, эффект суммации начинает уменьшаться; на этом этапе увеличение частоты возбуждения уже не усиливает напряжения мышцы, а лишь удерживает ее в сокращенном состоянии. Такая предельная реакция мышечного волокна называется тетанусом, и напряжение (не путать со степенью сокращения) мышечного волокна в это время примерно в четыре раза выше, чем во время одиночного сокращения (рис. 14). Поэтому для увеличения напряжения мышцы в процесс сокращения должно вовлекаться все больше и больше волокон.
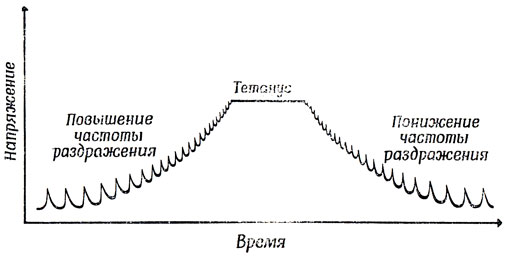
Рис. 14. Напряжение в одиночном мышечном волокне постоянной длины в ответ на увеличение и понижение частоты стимуляции. Одиночные сокращения, характерные для низкочастотного возбуждения, при достаточновысокой частоте приходящих импульсов уступают место длительному устойчивому (тетаническому) сокращению
При раздражении рецепторов увеличение частоты стимулов повышает генераторный потенциал. Если перейти к нейронам первого уровня, генерирующим спайки, то аксоны "разряжаются" каждый раз, когда возбуждают импульс, а затем должны вернуться в состояние, при котором порог сравним с потенциалом на входе. Стало быть, чем выше генераторный потенциал, тем короче будет этот период и, следовательно, тем выше будет частота импульсов. Поэтому на "входе" системы удобно приближенно описывать ситуацию так, как если бы интенсивность стимуляции можно было перекодировать в частоту импульсов. Но если уйти с периферии и обратиться к клеткам, ответственным за "вычисления", то нам придется проститься с такими простыми зависимостями и обратиться к взаимодействиям возбуждающих и тормозных клеток, где каждый тормозный импульс увеличивает разность между "текущим" потенциалом и его пороговым значением, а каждый возбуждающий уменьшает ее. Таким образом, по мере повышения частоты возбуждающих импульсов или уменьшения частоты тормозных частота импульсации будет возрастать.
Изложенные здесь данные создают основу, необходимую для изучения роли, которую играют нейроны в предварительной обработке зрительной информации у кошки и лягушки (разд. 2, 4). Однако некоторые читатели, возможно, захотят сначала представить себе место зрительной системы в общей анатомии мозга.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'