2.3. Отступление. Некоторые сведения о строении мозга
* (При первом чтении этот раздел можно пропустить.)
Мозг состоит из многих миллиардов нейронов. Для того чтобы выявить соотношение между функциями мозга в целом и функциями отдельных клеток, необходимо иметь некоторое представление о строении мозга. Рассматривая препараты мозга невооруженным глазом, можно увидеть, что мозг делится на ряд областей. Анализ с помощью различных микроскопических, биохимических и электрофизиологических методов позволяет выявить различия в свойствах, взаимосвязях и функциях разных областей и перейти к более тонкой "топографии" мозга.
Строению мозга, которое различается у разных видов, посвящена огромная литература. Анатомия нервной системы все еще находится в процессе изучения, а анатомия многих частей головного мозга столь же спорна, как и теории относительно функций мозга, излагаемые в этой книге! Однако, как я уже неоднократно подчеркивал, мы не можем надеяться понять, как работает мозг, не связывая его функции с его структурой. Поэтому здесь, а также в разд. 5.2 излагаются основные сведения по анатомии нервной системы. А поскольку многим читателям обилие незнакомых терминов, характерное для этих двух разделов, может показаться чрезмерным, книга написана так, что эти два раздела можно опустить без ущерба для понимания остальных. Я хотел бы также добавить, что мои познания в анатомии нервной системы весьма скромны, и поэтому мое изложение вряд ли имеет какие-либо особые достоинства, если не считать его краткости. Если в этом кратком очерке и есть что-то хорошее, то все это следует отнести за счет превосходных лекций У. Наута по анатомии нервной системы, которые он читал в 1966 г., в рамках программы подготовки к реализации исследовательского проекта по нейрофизиологии Массачусетского технологического института. Эти лекции впоследствии легли в основу одной из глав книги Наута и Картена "Общий очерк мозга позвоночных" [179].
Прежде всего следует сделать три замечания. Во-первых, в то время как некоторые области мозга имеют четкие границы (ни у кого не вызывает сомнений, какую часть мозга следует считать мозжечком), другие области постепенно переходят одна в другую. И относиться к границам между этими областями нужно так же, как к условным границам, разделяющим полоску бумаги, цвет которой постепенно переходит от белого к черному, на четыре области: "белую", "светло-серую", "темно-серую" и "черную", ясно сознавая, что нелепо спорить о том, где именно пройдет линия, отделяющая "белое" от "светло-серого". Во-вторых, точно так же как у некоторых людей бывает большая голова и короткое туловище, а у других - наоборот, так и относительное расположение и форма разных областей мозга сильно варьирует у разных индивидуумов. В-третьих, у разных групп животных строение мозга, несмотря на многие общие черты, сильно различается. Например, различия в предварительной обработке зрительной информации в сетчатке кошки и лягушки (вопрос, которым мы займемся в следующем разделе) ясно показывают, что одно лишь общее сходство некоторой части мозга лягушки с соответствующей частью мозга кошки еще не означает, что они выполняют одинаковые функции.
Человек вместе со всеми другими млекопитающими (а также птицами, рыбами, лягушками и крокодилами) относится к позвоночным. У него есть позвоночник - ряд полых костей (позвонков), расположенных вдоль спины и заполненных огромным количеством нейронов, образующих спинной мозг. Вся сенсорная информация от кожи, суставов и мышц туловища и конечностей поступает в спинной мозг, и клеточные тела всех мотонейронов, синапсы которых контролируют работу мышц туловища и конечностей, лежат в спинном мозгу. В отличие от этого информация, поступающая по черепно-мозговым нервам от рецепторов головы, приходит в центральную нервную систему главным образом через ствол мозга. Необходимо отметить, хотя здесь это нас интересовать не будет, что спинной мозг содержит также элементы автономной нервной системы, иннервирующей железы, гладкую мускулатуру стенок сосудов и внутренних органов, а также мышцы, поднимающие волосы.
У беспозвоночных строение нервной системы столь разнообразно и настолько сильно отличается от ее строения у позвоночных, что мы его касаться не будем. Мы займемся изучением нервной системы позвоночных и в данном разделе уделим основное внимание млекопитающим, и в частности человеку.
Прежде всего читателю придется вернуться к рис. 2, где показано расположение в голове человека основных отделов мозга, которые мы собираемся рассмотреть. На рис. 2, А хорошо видны структуры так называемого среднего мозга. Основное отличие головного мозга млекопитающих, и особенно человека, от мозга других видов состоит в резком увеличении новой коры, или неокортекса, которая у человека значительно превосходит по объему остальной мозг, что ясно видно на рис. 2, Б, где складки новой коры полностью закрывают средний мозг. Кора головного мозга человека представляет собой слой толщиной всего лишь в 60-100 нейронов, и для того чтобы в ограниченном пространстве черепа могли уместиться составляющие его несколько миллиардов нейронов, ему приходится образовывать множество складок. Как мы узнаем в разд. 5.2, такое резкое увеличение переднего мозга ведет к значительным изменениям структуры связей в стволе головного мозга и в спинном мозгу.
В каждом полушарии головного мозга принято различать четыре доли: лобную, височную, теменную и затылочную.
Складки коры головного мозга называют бороздами, а выступающую между двумя бороздами ткань - извилинами. Нервные пути, соединяющие различные области двух полушарий, называются комиссурами. Самая крупная из них - это мозолистое тело, осуществляющее связь между двумя полушариями.
Строение коры головного мозга неоднородно и различается в разных участках. Некоторые области коры называют сенсорными; в каждой из них в основном перерабатывается информация одной модальности; к ним относится не только соматосенсорная область (рис. 2,8), но также зрительная, слуховая и обонятельная (видная лишь на разрезе, поскольку она не выходит наружу) области, получающие информацию от дистантных рецепторов головы. Некоторые из нервных волокон, отходящих от моторной области коры головного мозга, идут через спинной мозг к мышцам и управляют движениями. Филогенетически соматосенсорная и моторная области неразрывно связаны друг с другом, и (по крайней мере) у человека не только в моторной области имеется сенсорное представительство (это и не удивительно, поскольку клетки, регулирующие движение, должны реагировать на соответствующие внешние стимулы), но более того, в соматосенсорной области имеются клетки, аксоны которых проецируются* (так же как и нейроны моторной области) на мотонейроны и вставочные нейроны спинного мозга, влияя таким образом на движение через посредство максимум двух промежуточных синапсов. В связи с этим область больших полушарий, примыкающих к центральной, или роландовой, борозде, принято называть просто сенсомоторной областью. Остальные области коры называют ассоциативными, но это неудачное наименование, отражающее ошибочное представление прошлого века, согласно которому функции этих областей заключались лишь в том, чтобы согласовывать, "ассоциировать" разнородные сенсорные данные и генерировать соответствующие команды, передаваемые в моторную область. Абсолютно неверное представление о том, что 90% мозга "не используется", распространенное среди дилетантов, возникло, вероятно, на той почве, что точные функции большей части этих "ассоциативных областей" до сих пор неизвестны.
* (Когда говорят, что данная область коры проецируется на другую, та это означает, что существуют нервные связи, ведущие из первой области во вторую.)
Мотонейроны, образующие синапсы на мышечных волокнах и управляющие их сокращениями, лежат в спинном, а не в головном мозгу. Особую группу эффекторных нейронов, не связанных с движением, составляют нейроны гипофиза. Гипофиз представляет собой важнейшую эндокринную железу. Он выделяет несколько гормонов, которые разносятся кровью по организму и управляют функцией других эндокринных желез. При этом важно помнить, что эндокринные железы образуют еще одну систему регуляции - гуморальную, дополняющую нервную систему. Это напоминание о разнообразии эффекторных механизмов нужно дополнить, заметив, что сенсорная поверхность организма не ограничивается его наружной поверхностью. Кроме внешних (экстерорецепторных) входов от наружной поверхности тела и таких органов чувств, как глаза, уши и нос, имеются сенсорные входы еще двух типов: во-первых, проприоцепторы, доставляющие сигналы обратной связи о движениях и ориентации туловища от мышц, суставов, сухожилий, отолитов и полукружных каналов, а во-вторых, интерорецепторы, передающие информацию от желез и внутренних органов.
Прежде чем перейти к объяснению других терминов, попытаемся снадбить анатомические структуры соответствующими функциональными коррелятами, рассмотрев два наиболее важных типа входов для рецепторов головы. В этой книге мы не будем касаться вкуса и обоняния. Отметим лишь, что обоняние, подобно зрению и слуху, обладает важной способностью регистрировать информацию об удаленных областях окружающей среды. Рис. 15 и 16 дают нам некоторое представление об общем строении зрительной и слуховой систем, занимающихся периферической обработкой реакции палочек и колбочек на распределение светового потока, отраженного от объектов внешнего мира, и волосковых клеток внутреннего уха, которые реагируют на колебания, вызванные звуковыми волнами, попадающими в ухо.
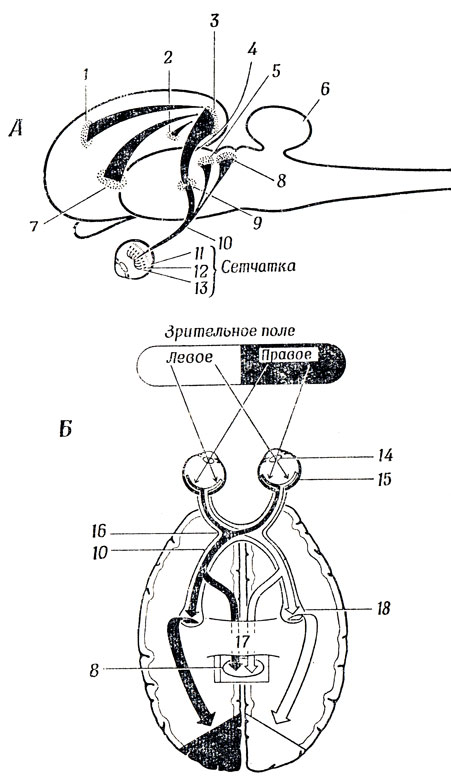
Рис. 15. Схема зрительной системы млекопитающих. А. Вид сбоку. Б. Горизонтальный разрез. Обратите внимание на наличие путей, ведущих как в новую кору (через латеральное коленчатое тело), так и в средний мозг (в претектальную область и верхний бугорок четверохолмия). 1 - фронтальная зрительная область; 2 - престриарная область; 3 - зрительная кора; 4 - зрительная радиация; 5 - претектальная область; 6 - мозжечок; 7 - нижневисочная кора; 8 - верхнее двухолмие (тектум); 9 - ядро латерального коленчатого тела; 10 - зрительный тракт; 11 - палочки и колбочки; 12 - биполярные клетки; 13 - ганглиозные клетки; 14 - хрусталик; 15 - сетчатка; 16 - хиазма; 17 - мозолистое тело; 18 - латеральное коленчатое тело
На рис. 15, А можно видеть, что потенциалы действия колбочек и палочек после предварительной обработки в двух слоях клеток сетчатки попадают в зрительный тракт (образуемый аксонами ганглиозных клеток), который делится на несколько путей. Два из них идут в средний мозг, к так называемому верхнему двухолмию, и к претектальной области (pretectum). Мы еще многократно будем возвращаться к верхнему двухолмию и к крыше мозга - его гомологу (т. е. структуре со сходным анатомическим расположением и сходным развитием в филогенезе) у лягушки (разд. 5.5 и 7.2), подчеркивая их роль в управлении взглядом животного. Третий пункт назначения зрительного тракта - и, надо сказать, самый важный у человека - расположен в таламусе и называется латеральным коленчатым телом. Здесь сигналы, идущие по зрительному тракту, проходят дополнительную предварительную обработку на пути к зрительной коре, расположенной в задней части головы. Зрительную кору называют также полем № 17 (по Бродману). Зрительная радиация, т. е. аксоны клеток латерального коленчатого тела, проецируется также на соседние поля № 18 и 19. Кроме двусторонней связи (не показанной на рис. 15) между зрительной корой и верхним двухолмием имеются также волокна, связывающие зрительную кору с другими областями коры, из которых на рисунке показаны лишь фронтальная зрительная область (контролирующая движение глаз) и нижневисочная кора (расположенная на внутренней поверхности височной доли).
В дополнение к виду сбоку на рис. 15,5 показана схема зрительной системы человека "сверху", на которой видны пути - от сетчатки к верхнему двухолмию и латеральному коленчатому телу. Здесь важно отметить, что зрительные тракты, идущие от двух глаз, перекрещиваются в области, которая называется зрительной хиазмой или перекрестом, таким образом, что аксоны, рецепторное поле которых находится в правом зрительном поле, поступают в левое полушарие мозга, а аксоны от левого зрительного поля - в правое.
Мы не воспринимаем правое и левое поля зрения по отдельности частично благодаря структурам среднего мозга, но в еще большей степени благодаря мозолистому телу - скоплению очень большого числа аксонов, по которым осуществляется связь между двумя полушариями. Захватывающие опыты Сперри и его сотрудников показывают, что если мозг "расщепить" на две части, перерезав мозолистое тело и другие комиссуры, то животное начинает вести себя так, как будто у него два самостоятельных представления о внешнем мире, соответствующих каждому полушарию. Человек с расщепленным мозгом может левым полушарием решить почитать газету, но в это же время правое полушарие может скомандовать положить газету на стол!
Переходя к рис. 16, мы получим наконец возможность ответить на животрепещущий вопрос: "А относительно чего латеральное коленчатое тело является латеральным?" Оказывается, оно латерально относительно медиального коленчатого тела, играющего для слуха такую же роль, какую латеральное коленчатое тело играет для зрения в том смысле, что это последняя "промежуточная станция" на пути к слуховой коре. Верхнее двухолмие называется так просто потому, что оно расположено над нижним двухолмием. Перекрестные входы в клетки верхней оливы позволяют им объединять информацию от обоих ушей, определяя пространственное расположение источника звука. Оказалось, что проекция нервных волокон на слуховую кору частично тонотипична, т. е. что место назначения этих волокон определяется тоном, или высотой звука, но обнаружение "детекторов мяуканья" и "детекторов щелчков" в слуховой коре головного мозга кошки усложнило картину. Слуховые рецепторы соединены также и с верхним двухолмием, в результате чего неожиданный шум может заставить зрительную систему заинтересоваться его источником. (На самом деле верхнее двухолмие связано с рецепторами всех органов чувств.) Подробным разбором слуховой системы мы в этой книге заниматься не будем и рекомендуем заинтересовавшемуся читателю обратиться к книге Уайтфилда [255] "Слуховые пути".
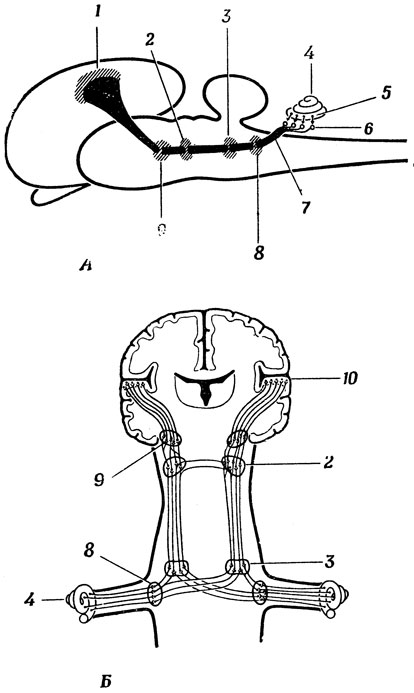
Рис. 16. Схема строения слуховой системы. А. Вид сбоку. Б. Фронтальный разрез, 1 - слуховая кора; 2 - нижнее двухолмие; 3 - верхняя олива; 4 - улитка; 5 - волосковые клетки; 6 - спиральный ганглий; 7 - слуховой нерв; 8 - кохлеарное ядро; 9 - медиальное коленчатое тело; 10 - слуховая кора в верхней височной извилине
В завершение этого раздела повторим еще раз то, что нам известно о разных отделах мозга, центральной нервной системы, используя для этого схему, приведенную на рис. 17 (рис. 18 позволит убедиться, сколь различно соотношение отдельных частей мозга у разных животных). В направлении от "головы" к "хвосту" нервная система делится на три основных раздела:
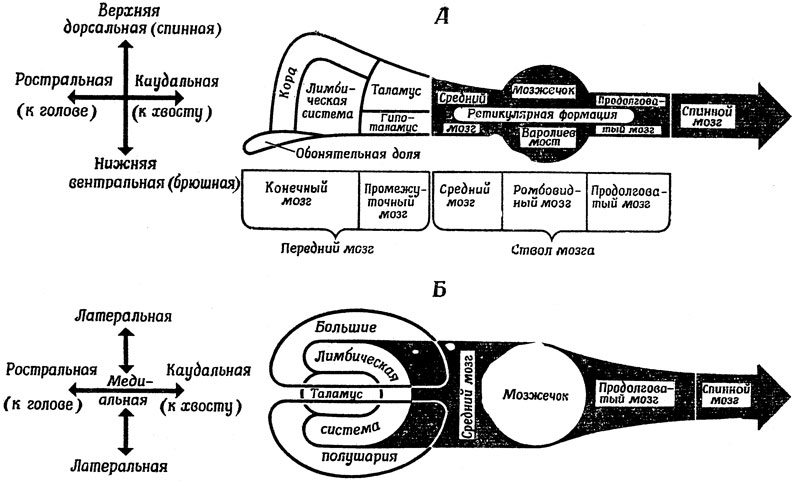
Рис. 17. Схема строения центральной нервной системы животного, не относящегося к млекопитающим. А. Вид сбоку. Б. Вид сверху
1) передний мозг, к которому относится конечный мозг и промежуточный мозг;
2) ствол мозга, к которому относится средний мозг, варолиев мост (но не мозжечок) и продолговатый мозг;
3) спинной мозг, где замыкаются рефлекторные дуги, а также проходят пути, по которым осуществляется связь сегментов спинного мозга между собой и с другими областями мозга.
Спинным мозгом мы займемся в разд. 5.1, а затем в разд. 5.2 рассмотрим устройство его основных двигательных механизмов. Остаток же этого раздела мы посвятим беглому обзору основных подразделов переднего мозга и ствола.
Конечный мозг, или телэнцефалон (от греч. телос - конец и энкефалос - мозг), образует у позвоночных самый передний отдел мозга. К нему относятся большие полушария, которые у млекопитающих резко увеличены в результате образования новой коры, или неокортекса (от лат. кортекс-кора). Полушария и доли мозга тесно связаны волокнами между собой и со средним мозгом многочисленными волокнами.
Лимбическая система лежит на границе между большими полушариями и стволом мозга. Эта система мультимодальна в том смысле, что в ней происходит объединение информации, поступающей от разных чувств; она играет особенно важную роль в поведении, направленном на выживание отдельной особи и вида в целом (т. е. в питании, чувстве опасности, агрессивности и размножении), причем ее реакции опосредуются через низшие центры промежуточного мозга. К лимбической системе относятся аммонов рог и миндалевидные ядра. Ее деятельность тесно связана с работой внутренних органов, физиологией эмоций и восприятием обонятельных стимулов (эту последнюю ее функцию иногда незаслуженно переоценивают, называя этот отдел обонятельным мозгом).
Обонятельные нейроны очень сходны с эпителиальными клетками, но образуют синапсы на обонятельных луковицах; отсюда сигналы направляются в обонятельную долю по обонятельному тракту.
На поперечном разрезе полушария головного мозга видно, что кора (т. е. поверхностный слой) состоит из серого вещества (т. е. в основном из тел клеток), а внутренние области мозга содержат как белое вещество (т. е. в основном аксоны), так и четко ограниченные участки серого вещества, расположенные у основания полушарий. Те из них, которые содержатся в переднем мозге, называются базальными ядрами и играют важную роль в управлении движением (разд. 5.2). Другие участки серого вещества - таламус и гипоталамус-расположены в промежуточном мозге, к рассмотрению которого мы сейчас и перейдем.
Промежуточный мозг, или диэнцефалон, состоит из таламуса и гипоталамуса. В некоторых областях таламуса происходит обработка сенсорной информации на пути к коре головного мозга; сюда же приходят аксоны из некоторых областей коры. Одна область дорсального таламуса преобразует моторные стимулы, посылаемые к мышцам, в результате чего сигналы, попадающие в нижележащие нервные клетки, вызывают организованную реакцию. Медиальное и латеральное коленчатые телаг о роли которых в обработке зрительной и слуховой информация мы уже говорили, также относятся к таламусу.
Гипоталамус контролирует биохимические процессы, происходящие в организме; он регулирует солевой обмен, кровяное давление, дыхательные процессы, температуру тела, водный баланс, чувство голода и насыщения, эндокринную систему и вкупе с лимбической системой играет решающую роль в эмоциональном и мотивационном поведении человека.
Перейдем теперь к стволу мозга и мозжечку.
Средний мозг, или мезэнцефалон, регулирует зрительные и слуховые рефлексы, а также общедвигательные рефлексы. Он содержит многочисленные восходящие и нисходящие пути, имеющие отношение к соматосенсорной и примитивной моторной системам. Здесь находится также так называемая ретикулярная формация с комплексами нейронов, обеспечивающими компенсацию силы тяжести, гомеостаз и чередование сна и бодрствования.
К заднему мозгу относятся варолиев мост и мозжечок, а также многие восходящие и нисходящие пути. Отсюда отходят волокна к коре мозжечка, идущие контрлатерально* и передающие информацию о характере импульсов, посылаемых к нижележащим мотонейронам спинного мозга. Варолиев мост содержит также ядра черепно-мозговых нервов, ретикулярные ядра и восходящие (сенсорные) и нисходящие (примитивные двигательные) системы волокон. Мозжечок посылает сигналы как в высшие, так и в низшие центры, поддерживает тонус определенных групп мышц и осуществляет регуляцию мышц-синергистов и антагонистов, что обеспечивает координацию движений. Это, возможно, достигается благодаря связям с ретикулярной формацией среднего мозга и проекциям на сенсомоторную кору, идущим через таламус; через эти проекции мозжечок оказывает влияние на регуляцию последовательности двигательных актов, осуществляемую корой.
* (Волокна, начинающиеся по одну сторону срединной плоскости и проецирующиеся на другую (противоположную) сторону тела. Аналогично ипсилатеральными называют волокна, остающиеся на всем своем протяжении по одну сторону срединной плоскости.)
Продолговатый мозг является как бы продолжением спинного мозга в черепе. Он играет важную роль в регуляции сердечно-сосудистой системы, дыхания, пищеварения и в поддержании равновесия. Системы длинных нисходящих и восходящих путей проходят и через продолговатый мозг. Пучки восходящих сенсорных волокон, берущие частично свое начало в спинном мозге, передают информацию о взаиморасположении различных частей тела и мышечном тонусе в мозжечок.
Заканчивая этот беглый обзор, напомним еще раз, что разные животные могут отличаться по строению мозга не в меньшей степени, чем они отличаются по внешнему виду. Некоторое представление о разнообразии строения мозга дает рис. 18, в еще большей степени в этом можно убедиться при сравнении зрительных систем лягушки и кошки, которому посвящен следующий раздел.
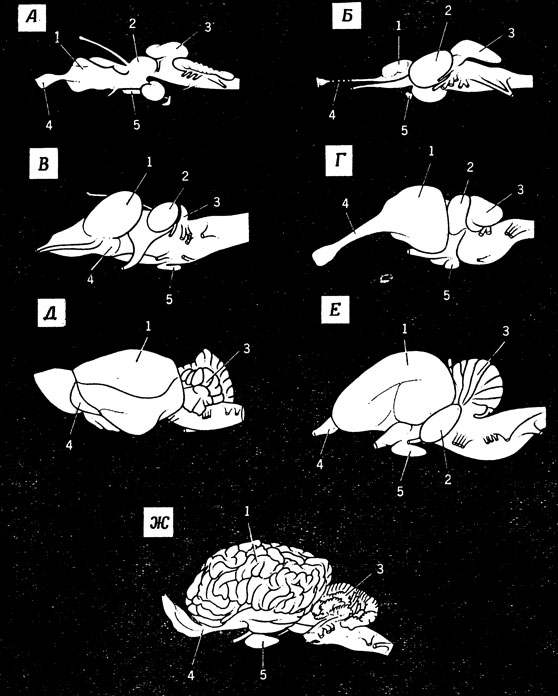
Рис. 18. Схемы строения мозга у разных животных (вид сбоку). А. Акула. Б. Треска. В. Лягушка. Г. Крокодил. Д. Примитивное млекопитающее. Е. Гусь. Ж. Лошадь. 1 - головной мозг; 2 - зрительная доля; 3 - мозжечок; 4 - обонятельная доля; 5 - гипофиз
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'