5.1. Обратная связь и спинной мозг
При изучении нервных механизмов моторной деятельности очень важно понять центральную роль спинного мозга в организации движения конечностей и их координации, который обеспечивает компенсацию непрерывно меняющейся внешней обстановки и осуществляет коррекцию позы; это дает возможность организму сохранять равновесие при сложной двигательной активности. В этом разделе мы познакомимся с некоторыми контурами обратной связи спинного мозга, а в следующем опишем в общих чертах механизмы управления "высшего уровня", связанные с поведением.
Спинной мозг (рис. 76) расположен в позвоночнике. На поперечном сечении спинного мозга можно отчетливо различить серое вещество (тела нервных клеток и дендриты) и белое вещество (в основном аксоны). Входные и выходные каналы четко разделены: тела первичных сенсорных клеток (клеток, дендриты которых "специализированы" на получении сенсорной информации) сосредоточены в ганглиях задних корешков, а их аксоны попадают в задние рога спинного мозга по задним корешкам. Тела мотонейронов расположены в передних рогах спинного мозга, а их аксоны идут в составе передних корешков. Передний и задний корешки объединяются, образуя нервный ствол. Все тело животного разделено на ряд сегментов, каждый из которых иннервируется нервами, отходящими от соответствующего сегмента спинного мозга.
На рис. 76 видно, что спинной мозг состоит в основном из белого вещества. Здесь проходит множество восходящих и нисходящих нервных волокон, связывающих различные сегменты спинного мозга между собой, а также спинной мозг с головным. Серое вещество разделяет белое вещество спинного мозга на четыре тяжа, получивших название столбов: два боковых (или латеральных), один задний (дорсальный) и один передний (вентральный).
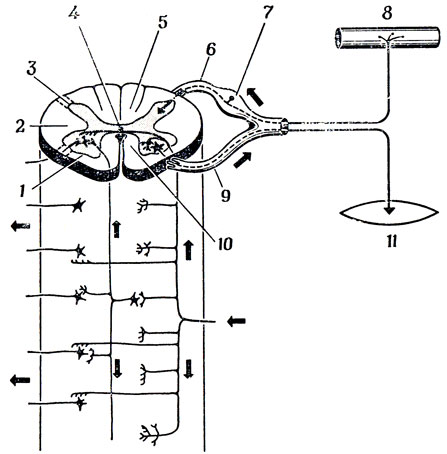
Рис. 76. Схема поперечного сечения спинного мозга. Сенсорная информация (в том числе и сигналы обратной связи) поступает по задним корешкам, а управляющие сигналы к эффекторам уходят по передним корешкам. Нервная сеть, изображенная под поперечным сечением, должна напоминать о богатстве восходящих и нисходящих путей, идущих по спинному мозгу. 1 - передний рог; 2 - боковой столб; 3 - задний рог; 4 - центральный канал; 5 - задний столб; 6 - задний корешок; 7 - ганглий заднего корешка; 8 - мышечные веретена и сенсорная поверхность; 9 - передний корешок; 10 - передний столб: 11 - мышцы-эффекторы
О значении путей, приходящих в спинной мозг извне, свидетельствует тот факт, что движение мышцы может возникнуть не только в ответ на что-то увиденное или услышанное, но и может быть вызвано мыслью или прикосновением. Другими словами, мотонейрон может реагировать на входные сигналы любой сенсорной модальности или даже на смесь таких сигналов, причем они могут смешиваться даже очень далеко от периферии. В этом разделе мы сосредоточим свое внимание на непосредственных связях между сенсорными клетками и мотонейронами (хотя таких связей относительно мало), к которым относятся и моносинаптические рефлекторные дуги, где между рецептором мышцы и мотонейроном имеется всего один промежуточный синапс, и лишь весьма кратко упомянем о связях между различными сегментами спинного мозга, обеспечивающими координацию движений конечностей и автоматическое поддержание вертикального положения тела.
В таком сжатом описании, естественно, нет возможности уделить должное внимание всем деталям строения спинного мозга, которые должна учитывать любая действительно глубокая теория движения животных. Мы же преследуем здесь более ограниченную цель - дать общее представление о том, к чему привел бы подобный детальный анализ.
То, что мы привыкли называть мышцей, на самом деле состоит из тысяч отдельных волокон. Каждый мотонейрон, тело которого расположено в спинном мозгу, образует синапсы на нескольких из этих мышечных волокон и управляет ими. Те мотонейроны, которые управляют тонкими движениями, например движениями пальцев у приматов, воздействуют на очень небольшое число волокон каждый. В других случаях, например в плече, один мотонейрон может управлять тысячами мышечных волокон. Волокна группируются в пучки, из которых в свою очередь образуются мышцы, причем в отдельных мышцах число волокон достигает нескольких миллионов. Волокна одной двигательной единицы (т. е. волокна, иннервируемые одним мотонейроном) входят в состав разных пучков, благодаря чему действие каждой двигательной единицы распространяется на всю мышцу., Существует два пути повышения уровня развиваемого мышцей усилия: один из них, играющий более важную роль, основан на увеличении числа волокон, включающихся в процесс сокращения в результате синаптической активации; другой - повышение активации отдельного мышечного волокна - приводит к гораздо менее значительному увеличению активности. По мере сокращения мышцы в процесс включаются все новые группы волокон; сначала относительно мелкие волокна, затем покрупнее, затем еще более крупные, причем на каждом этапе относительное приращение усилия примерно одинаково, что способствует плавности движений.
Из разд. 2.2 мы уже знаем, что можно приказать мышце перестать сокращаться, но нельзя непосредственно заставить ее расслабиться, и поэтому противоположными движениями управляют пары мышц-антагонистов (рис. 77).
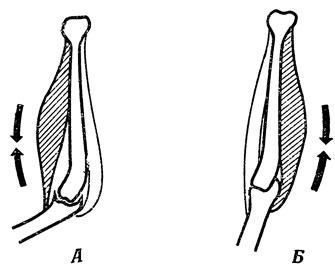
Рис. 77. Пара мышц-антагонистов. Активное сокращение мышцы-сгибателя вызывает пассивное расслабление ее антагониста - мышцы-разгибателя (А) и наоборот (Б). Сокращаясь, сгибатель уменьшает суставной угол (А), а разгибатель - увеличивает его (Б)
Однако в управлении движением в таких суставах, как плечевой, имеющий более одной степени свободы, участвует много мышц, и здесь нет особого смысла выделять пары мышц-антагонистов (см. рис. 36).
В большинство естественных движений вовлекаются сразу несколько суставов. Например, для того чтобы с максимальной силой ухватиться за что-нибудь рукой, кисть должна быть фиксирована в состоянии разгибания. Этим занимаются специальные мышцы, которые можно назвать фиксаторами. Мышцы, работающие одновременно и поддерживающие мышцу, осуществляющую основное движение, называются синергистами. Например, если мы пытаемся поднять что-нибудь одним пальцем, в работу включается сразу много мышц. Если развиваемое при этом усилие недостаточно, то в действие вступают и мышцы кисти; если и этого мало - включаются мышцы предплечья, а на этом этапе начинают действовать и многие другие мышцы, расположенные по всему телу и обеспечивающие поддержание позы.
Спинной мозг осуществляет многие важные рефлексы. Например, у спинального животного (т. е. у животного, у которого перерезан спинной мозг на уровне верхних сегментов, так что головной мозг не участвует в управлении движениями) наблюдается чесательный рефлекс: оно может дергать ногой, стараясь почесать раздражаемый участок тела. При этом движения в суставах происходят одновременно (синергия). Те же общие реакции наблюдаются и у интактного животного (с ненарушенной нервной системой), но в этом случае движения в разных суставах могут изменяться. Таким образом, помимо замкнутого спинального контура низшего уровня, обеспечивающего, как мы увидим ниже, рефлекторные движения, независимо от величины нагрузки, без помощи высших центров, в этих грубых макродвижениях могут участвовать и высшие центры, обеспечивающие их тонкую настройку. Даже после перерезки спинного мозга кое-какая информация может передаваться от головы к телу. Если повернуть голову, то кожа на шее передаст сведения об этом телу, что, возможно, вызовет естественные позные рефлексы. К тому же движения головы изменяют положение центра тяжести. Тем не менее можно утверждать, что при перерезке спинного мозга поток информации от головы к телу очень резко сокращается и что остающейся информации явно недостаточно для управления упомянутыми рефлексами.
В связи с этим мы будем считать, что роль головного мозга состоит в упорядочении выбора мышц-синергистов, последовательности их включения в работу и подавления нежелательных составляющих. Такая организация управления, вероятно, эффективна для большинства задач, но, естественно, она ограничивает репертуар возможных движений. (Например, становится трудна поглаживать себя одной рукой по животу, одновременно похлопывая другой рукой по голове. А то, что человеку это удается, свидетельствует о новом уровне приспосабливаемости, достигнутом за счет развития новой коры головного мозга.) Таким образом, главная наша задача заключается в том, чтобы понять, как организм целенаправленно выбирает группу мышц-синергистов, изменяет и координирует их совместную работу.
В разд. 3.3 мы уже убедились в том, что обратная связь (рис. 78), т. е. возхможность сравнивать реальный результат с "желаемым", должна играть важную роль в процессах управления как в организме, так и у робота. Подобно тому как управляя автомобилем, мы для поддержания постоянной скорости должны нажимать на тормоз, если дорога идет под гору и на газ, если дорога пошла вверх, так и в нервной системе неразумно было бы ожидать, что для обеспечения постоянства реакции от высших центров непрерывно поступают какие-то постоянные команды. Напротив, для плавных координированных действий, вероятно, необходимы весьма сложные и разнообразные процессы управления. Посмотрим, в частности, как локальные контуры обратной связи в спинном мозгу обеспечивают требуемое поведение в условиях меняющейся нагрузки.
Обратная связь используется, например, в нервной системе для поддержания относительно постоянной длины мышцы при небольших изменениях нагрузки. Для того чтобы понять, каким образом ЦНС получает сигналы обратной связи для компенсации такого рода, возьмем кусок резинки, длина которой в нерастянутом состоянии равна l0. Закрепим оба конца этой резинки зажимами, которые можно разводить относительно друг друга на разные расстояния l (рис. 79). Выберем на резинке две произвольные точки A и B, расстояние между которыми при нерастянутом состоянии резинки равно d0; тогда до тех пор, пока расстояние между зажимами будет не больше l0, т. е. резинка будет оставаться нерастянутой, расстояние между этими точками будет оставаться равным d0. Но если зажимы развести на большее расстояние, то резинка растянется и расстояние между точками A и B возрастет пропорционально увеличенной длины всей резинки. Это новое расстояние d между A и B можно подсчитать по формуле d=(l/l0)•d0 если резинка растянута. А это позволяет нам определить l косвенно, измерив расстояние между точками А и В и восстановив по нему значение l по формуле l=l0(d/d0), работающей всякий раз, когда l больше, чем l0.
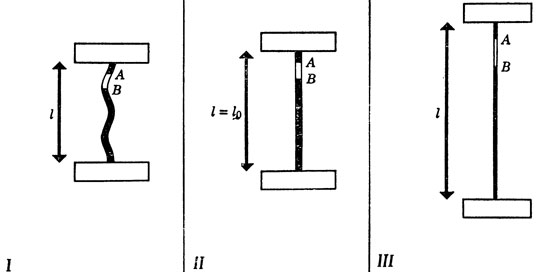
Рис. 79. Длина участка AB пропорциональна расстоянию l между двумя зажимами при условии, что это расстояние не меньше длины l0 резинки в нерастянутом состоянии. I. l<l0 - резинка не растянута и 'болтается'. II. l=l0 - резинка не растянута, но распрямлена. III. l>l0 - резинка растянута
Описанная схема отражает принцип получения нервной системой информации о длине мышцы. В любой мышце можно увидеть не только крупные "рабочие" мышечные волокна, называемые экстрафузальными, сокращение которых изменяет взаиморасположение костей, к которым они прикреплены, но и мелкие интрафузалъные волокна, на которых расположены окончания особых рецепторов, мышечных веретен; интенсивность разряда мышечных веретен сообщает нервной системе о степени растяжения мышечных волокон в соответствующей области па тому же самому принципу, как в случае резинки по длине "участка AB" можно было определить длину резинки в целом.
Таким образом, контур обратной связи, используемый ЦНС Для управления длиной мышцы, в основных чертах выглядит так, как показано на рис. 80; для простоты мы изобразили на схеме один мотонейрон, одно экстрафузальное и одно интрафузальное волокно, тогда как на самом деле в каждой крупной мышце имеются целые популяции таких элементов. Будем считать, что верхняя точка закреплена неподвижно, а нижняя может перемещаться вверх и вниз при различных условиях нагрузки и сокращения мышцы. На нашей схеме α-мотонейрон (т. е. мотонейрон, управляющий "рабочими" экстрафузальными волокнами) получает входные сигналы двух типов: "внешние" стимулы (комбинированный эффект сигналов "высшего управления", взаимодействия нескольких конечностей и т. п.) и сигналы "обратной связи", приходящие от мышечных веретен. До тех пор пока внешний стимул остается достаточно сильным для того, чтобы "рабочие" волокна могли поддерживать общее сокращение мышцы, не вызывая растяжения мышечного веретена, рецепторная иннервация почти ничего не добавляет к этому "внешнему" стимулу. Предположим, однако, что эти "внешние" стимулы таковы, что они поддерживают мышцу в состоянии, соответствующем состоянию интрафузального волокна на грани растяжения. Если подвесить затем к нижнему зажиму дополнительный груз, то произойдет следующее:
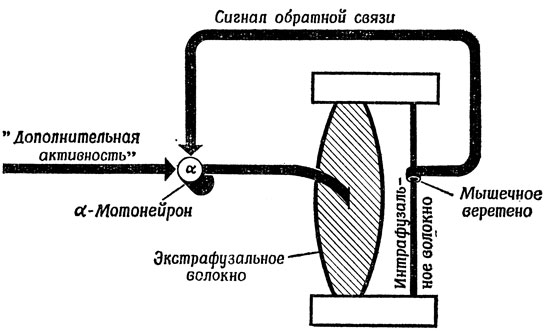
Рис. 80. Система с обратной связью для регулирования длины мышцы, построенная на принципе, представленном на рис. 79. Мышечное веретено контролирует степень растяженгя интрафузального волокна относительно длины покоя, а получаемый сигнал используется для увеличения активности α-мотонейрона, что в свою очередь вызывает сокращение экстрафузального волокна до тех пор, пока это растяжение не компенсируется
1) дополнительный вес вызовет дополнительное растяжение мышцы;
2) в результате этого растянется и мышечное веретено, которое начнет генерировать сигналы обратной связи;
3) это приведет к повышению суммарного уровня возбуждения на входе, а значит, и на выходе α-мотонейрона;
4) вследствие этого мышца дополнительно сократится и с течением времени по крайней мере частично вернется к своей, прежней длине.
У этой схемы организации управления есть две отличительные особенности.
I. Можно сокращать мышцу до любой длины, меньшей чем длина покоя интрафузального волокна, вызывая достаточно интенсивный разряд α-мотонейрона, но, к сожалению, размер необходимого возбуждения варьирует в зависимости от нагрузки.
II. Можно обеспечить сохранение длины мышцы при минимальной чувствительности к изменениям нагрузки, но, к сожалению, это справедливо лишь в том случае, когда длина мышцы равна длине покоя соответствующего интрафузального волокна.
Догадливый читатель, возможно, уже нашел выход из этих затруднений: нужно изменять длину покоя интрафузального волокна. Возвращаясь к нашему примеру с резинкой (рис. 79), представим себе, что мы завязываем узлы в средней части резинки (рис. 81, А). Нервная система достигает подобного эффекта благодаря тому, что в ней, кроме а-мотонейронов, управляющих сокращением "рабочих" экетрафузальных волокон, имеется множество у-мотонейронов (рис. 81, Б); умотонейроны образуют синапсы в средней части интрафузальных волокон и могут вызывать их сокращения, что приводит к уменьшению эффективной длины покоя. (Напомним, что все эти мотонейроны расположены в спинном мозгу, и их аксонам приходится передавать возбуждение на большие расстояния, прежде чем они достигнут мышечных волокон.)
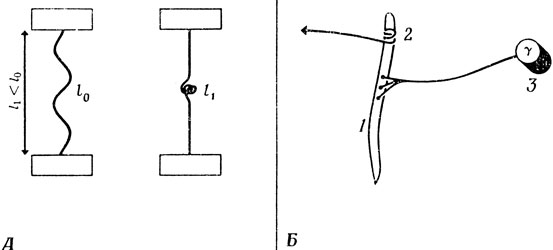
Рис. 81. Длину покоя резинки можно уменьшить, завязав на ней несколько узелков (A, справа), а длину покоя интрафузального волокна можно уменьшить, вызвав его сокращение, управляемое γ-мотонейроном. 1 - область, сокращающаяся при активации мотонейрона; 2 - мышечные веретена; 3 - γ-мотонейроны
Итак, если нам надо изменить длину мышцы, мы можем соответствующим образом возбудить α-мотонейроны, обеспечивающие требуемое сокращение мышцы в условиях минимальной нагрузки, одновременно возбуждая γ-мотонейроны, с тем чтобы вывести интрафузальные волокна на новый уровень растяжения а обеспечить необходимые сигналы обратной связи относительно новой длины волокна. Таким образом, α-активация быстро выводит длину мышцы в "требуемую окрестность", а γ-активация осуществляет "тонкую настройку" длины, подгоняя активность α-мотонейронов к причудливым изменениям внешней нагрузки.
В действительности же дело обстоит еще сложнее. Например, как показывает рис. 82, в рассматриваемой системе есть еще один источник сигналов обратной связи - так называемые тельца Гольджи, находящиеся в сухожилиях; при мышечном сокращении эти тельца растягиваются и в них возникают нервные импульсы, поступающие в нервные центры и сообщающие о размерах усилий, развивающихся в сухожилиях. Кроме мышечных веретен, реагирующих на изменение длины мышцы, о которых здесь говорилось, существуют и другие рецепторы, контролирующие по типу обратной связи скорость растяжения, участвуя тем самым в обеспечении плавности происходящих движений (в отличие от стабилизации заданной позы). И наконец, необходимо отметить все еще нерешенную проблему распределения управления движением между α- и γ-системами. Но несмотря на все эти нерешенные вопросы, мы по крайней мере начинаем понимать, как α-система может обеспечить быструю и приблизительно верную реакцию, после чего в игру вступает обратная связь, освобождающая нервную систему от необходимости учитывать в своих расчетах незначительные изменения периферических условий. Возвращаясь к нашим рассуждениям в разд. 3.3, испытываешь большое искушение считать, что α-система обеспечивает баллистическую, а α-система - следящую составляющую комбинированной баллистико-следящей стратегии, которая позволяет быстро переводить систему в новое состояние, не теряя в то же время преимуществ, которые дает компенсация ошибок с помощью сигналов обратной связи.
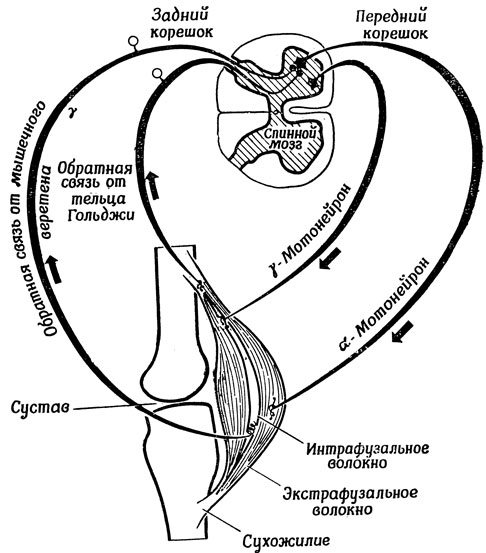
Рис. 82. Менее схематичное изображение механизмов, представленных на рис. 80 и 81, показывающее взаиморасположение мышц и костей, а также контуры управления, проходящие через спинной мозг
Получив некоторое представление о роли обратной связи в упрощении задач управления изолированной мышцей для "высших центров", займемся теми следствиями, которые вытекают из наших представлений о том, что движениями в каждом суставе управляет по меньшей мере пара мышц. Рассмотрим пару сгибатель - разгибатель, схематически изображенную на рис. 83. Совершенно ясно, что угол в этом суставе зависит от того, насколько сокращение сгибателя превосходит по силе сокращение разгибателя. Не менее ясно, что было бы расточительством сильно сокращать обе мышцы, чтобы добиться разницы, которую можно получить и при незначительных сокращениях тех же мышц. Поэтому, если нервной системе нужно, например, уменьшить угол в суставе, она должна не только увеличить сокращение сгибателя, но и уменьшить сокращение разгибателя. На рис. 83 представлена схема с обратной связью, автоматически осуществляющая такие координированные действия. На этой схеме рецептор мышечного веретена не только возбуждает α-мотонейрон своей собственной мышцы, но и понижает возбудимость, обычно (хотя, возможно, и не всегда; см. [224]), через посредство тормозного вставочного нейрона, α-мотонейрона мышцы-антагониста. Поэтому выходной сигнал мышечного веретена не только увеличивает сокращение мышцы-агониста, но и уменьшает сокращение мышцы-антагониста, обеспечивая экономию мышечных усилий при выполнении заданного движения.
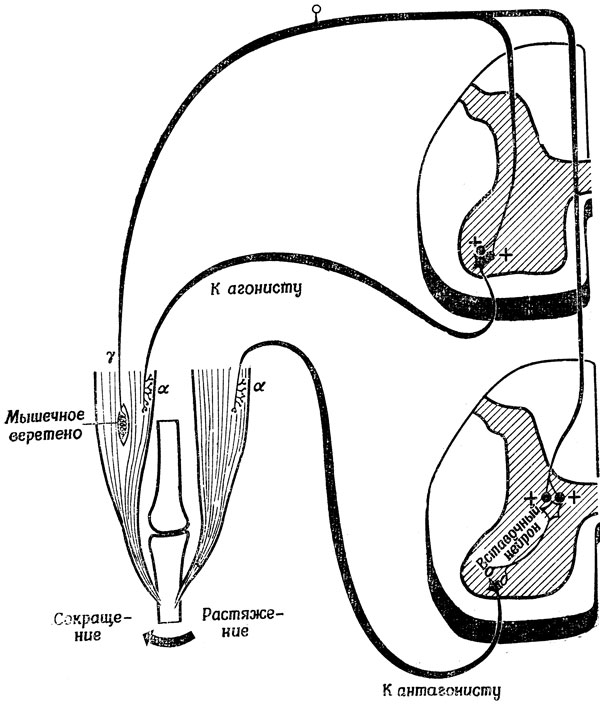
Рис. 83. Схема, дополняющая рис. 82. Обратная связь от мышечного веретена мышцы-агониста не только повышает уровень возбуждения α-мотонейрона, управляющего сокращением этой мышцы, но кроме того, через посредство тормозного вставочного нейрона понижает уровень возбуждения α-мотонейрона, управляющего сокращением мышцы-антагониста
Приведенные выше соображения позволяют понять основные достоинства сочетания иерархической организации управления с локальными контурами обратной связи, "уменьшающими загрузку высших уровней вычислительными задачами". Мы могли бы продолжить начатое здесь исследование локальных контуров управления, показав, как взаимосвязаны нейроны, принимающие участие в управлении различными суставами одной и той же конечности, каким образом две конечности одной пары связаны внутрисегментарными рефлексами, получившими свое название, потому что они происходят в пределах одного сегмента спинного мозга, и как на этой основе достраиваются межсегментарные рефлексы, объединяющие действия различных сегментов спинного мозга и обеспечивающие плавные автоматические коррекции позы или последовательность шагов во время локомоции. Однако, вместо того чтобы подробно разбирать здесь все эти связи (если бы мы даже и решились на это, многие наши рассуждения оказались бы спекулятивными, ибо спинной мозг - настолько сложная система, что в нем до сих пор еще немало загадок для нейрофизиолога), мы попытаемся дать читателю возможность почувствовать всю их сложность на примере процесса ходьбы (разд. 5.3). Некоторые читатели, быть может, захотят сначала ознакомиться с общим очерком анатомии некоторых отделов головного мозга, участвующих в управлении движением.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'