5.2. Еще одно отступление. Нейроанатомия двигательной системы
Цель этого раздела (его можно опустить без ущерба для понимания дальнейшего материала) состоит в том, чтобы на основе данных, изложенных в разд. 5.1, наметить схему различных спинномозговых рефлекторных путей, для которых сенсорная информация поступает в спинной мозг по задним корешкам, а "команды" мотонейронов покидают его по передним корешкам. При этом может оказаться полезным вспомнить терминологию, введенную в разд. 2.3.
Прежде всего познакомимся с общей организацией двигательной системы, схематически представленной на рис. 84. Эта схема основана на рисунках Наута, о лекциях которого уже упоминалось на разд. 2.3. Рассмотрим сначала белые блоки и связи, показанные сплошными линиями. Помимо "локальных каналов", являющихся одновременно как внутри-, так и межсегментарными, у позвоночных имеются еще два важных канала. Четвертый канал - "новый" спино-таламический тракт - имеется только у млекопитающих; его мы рассмотрим позднее. Второй канал, восходящий от вторичных сенсорных клеток к мозжечку, расположенному позади ствола мозга, обеспечивает мозжечковый контроль. Третий канал образован древним лемнисковым трактом и встречается у животных, находящихся на более низкой стадии развития, чем млекопитающие (по причинам, которые станут ясными в разд. 5.3, этот контур на схеме не показан). Лемнисковый тракт представляет собой "древнюю" часть спиноталамического тракта, состоящего из двух филогенетически различных компонентов, которые несут различные функции и оканчиваются в разных центрах. Этот тракт идет от сенсорных ядер в передний мозг через клетки таламуса. [Перерезка спиноталамического тракта вызывает по крайней мере временное исчезновение чувства боли (в связи с этим его иногда называют "болевым трактом"). Полезная роль боли состоит в том, что она вызывает активацию ретикулярной формации (о чем речь пойдет ниже).]
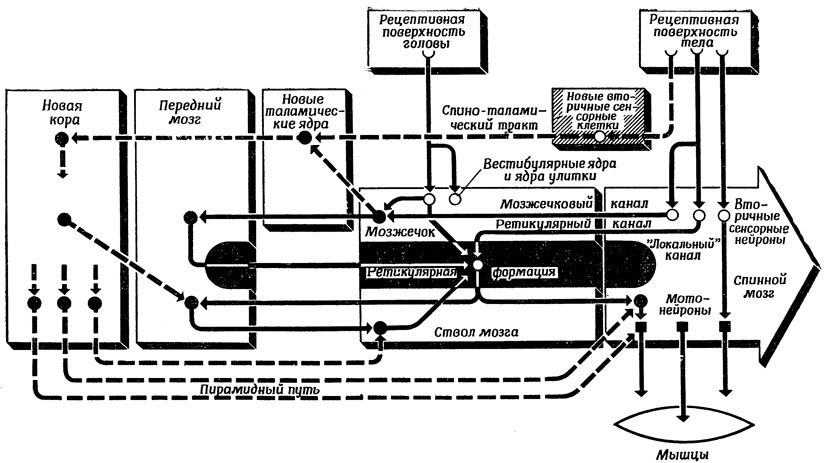
Рис. 84. Изменения строения и функций нервной системы позвоночных в процессе эволюции. Заштрихованные блоки и пунктирные линии - нервные структуры и системы связи позвоночных более низкого уровня, чем млекопитающие. Белые блоки и сплошные линии - структуры и системы связи млекопитающих
Даже у человека лишь примерно одна десятая волокон спино-таламического тракта идет непосредственно в таламус, а остальные "отклоняются" и уходят в ретикулярную формацию (РФ). РФ образована нейронами ствола и спинного мозга (но не переднего мозга), которые не специализированы по сенсорным модальностям. (Ретикулярная формация может служить классическим примером области со спорными границами. Предложенное выше определение является, вероятно, наиболее широким.)
Клетки РФ часто называют "неспецифическими" клетками, поскольку они способны воспринимать сенсорные стимулы разной модальности. Однако не следует считать, что у нейронов РФ нет специфических функций. В том, что зачастую кажется нам хаотичным, далеко не всегда отсутствует всякий порядок. В пределах РФ наблюдается временная миграция функций, проявляющаяся в том, что, по-видимому, одна и та же клетка в разные моменты времени может настраиваться на прием совершенно различных стимулов. Это не должно нас удивлять, если только мы не слишком стремимся переносить то, что нам известно о периферических проекциях, на весьма сложным образом взаимосвязанные центральные клетки. Не следует также считать, что подобная миграция функций свидетельствует о неограниченных возможностях каждого нейрона. В любом таком изменении нужно уметь увидеть заложенные в нем степени свободы, так же как и в позных рефлексах, которые вызывают удивление лишь до тех пор, пока мы не поймем, что все происходящее строго предопределено. Такое отсутствие монополии неизбежно, коль скоро речь идет об области, в задачу которой входит управление теми или иными функциями, причем характер управления зависит от многих модальностей: мы можем поднять ногу, потому что наступили на гвоздь или увидели перед собой ступеньку, или услышали приказ и т. п.
РФ обладает чрезвычайно широким диапазоном регуляторных функций. У отдаленных предков человека она, возможно, представляла собой высший уровень нервной регуляции. РФ и до сих пор играет большую роль в регуляции "позы" и "устойчивости". Вместе с локальными каналами спинного мозга она осуществляет поддержание позы в гравитационном поле, т. е. обеспечивает устойчивость организма во внешнем пространстве РФ проникает также в гипоталамус, вместе с которым она обеспечивает гомеостаз организма, т. е. примерное постоянство внутренней среды (содержание в крови углекислоты, сахара и т. п.), функционирование механизма дыхания и регуляцию кровяного давления. РФ несет и свои особые функции, поддерживая различные "типы" поведения организма, и в первую очередь - чередование периодов сна и бодрствования, а также регулируя переключение с одного режима на другой. Из всех функций РФ мы наиболее тщательно познакомимся именно с системой управления "типами" поведения, модель которой описана в разд. 7.1.
Перейдем теперь к заштрихованным блокам и пунктирным связям на рис. 84. Новая кора (неокортекс) полушарий головного мозга - характерный признак млекопитающих, "скачкообразно" возникший в процессе эволюции. Чем больше развита новая кора, тем на более длительные сроки может животное "планировать" свою деятельность. Отделы мозга, начиная от среднего мозга и ниже, заняты тем, что происходит "здесь и в данный момент". Даже шимпанзе неспособны подобно человеку предвидеть будущее и учитывать это при планировании своих действий. Долгосрочному планированию способствуют также сложные социальные взаимодействия, ставшие возможными благодаря тому, что различные области коры головного мозга человека приспособлены к управлению его речевой деятельностью.
Вместе с новой корой появились и новые контуры управления. Образовались новые ядра (группы клеток) таламуса, на которых находятся синапсы спино-таламического тракта, несущего в новую кору информацию, очищенную от "примесей" эффектов древних путей, проходящих через РФ. Появился также новый прямой двигательный тракт, названный пирамидным, передающий команды непосредственно от новой коры в спинной мозг. У приматов, способных производить весьма тонкие манипуляции (может, их следовало бы называть "педипуляциями", поскольку они производятся ногами?), имеются даже прямые связи между новой корой и отдельными мотонейронами.
Эти новые структуры существуют не обособленно от более старых, а скорее расширяют и модифицируют функции старых структур. С подобной схемой модификации функций, вызванной появлением новой коры, мы познакомимся в разд. 6.3, где будут намечены естественные возможности соматотопического подключения зрительной коры к верхнему двухолмию, обеспечивающегр большее совершенство зрительного поведения млекопитающих по сравнению с более примитивными позвоночными. Пирамидный путь также имеет тенденцию идти в обход более древних двигательных путей, но лишь 10% его волокон идут непосредственно к мотонейронам. Возможно, что это объясняется как тем, что регуляция движения носит в основном иерархический характер, т. е. высшие центры определяют структуру возбуждения целых групп мотонейронов, а не каждого из них, так и тем, что большая часть исходящих сигналов несет прогноз того, что должны делать такие группы эффекторов, и, следовательно, направляются в относительно высокорасположенные спинномозговые центры, где осуществляется контроль сигналов обратной связи от низших центров и с периферии. Затем такие "корреляционные центры" получают возможность "доложить" головному мозгу о том, насколько происходящие движения "соответствуют плану". (Все это весьма напоминает идею внутренней обратной связи [69].) Каждой структуре возбуждения высших центров, ответственной за инициацию движения, соответствует определенная структура возбуждения сенсорных центров, и каждое осуществленное движение вызывает изменение структуры сенсорного входа. Таким образом, между сенсорной и моторной системами существуют как внутренние, так и внешние взаимосвязи. Занятно, что в литературе "новые" пирамидные пути получили название "классических". Это объясняется тем, что клиницисты-невропатологи разобрались в них гораздо раньше, чем удалось распутать филогенетически более древние пути, идущие от РФ. Помимо этих совершенно новых путей, неоспиноталамического (или классического лемнискового восходящего) сенсорного тракта и пирамидного (классического нисходящего) двигательного тракта, имеются также новые пути, надстроенные над старыми. Это параллельный экстралемнисковый восходящий сенсорный тракт, надстроенный на филогенетически более древнем лемнисковом тракте; оба пути объединяются на уровне среднего мозга, откуда они разветвляются, достигая различных областей коры и проходя через РФ ствола мозга; это и параллельный экстрапирамидный двигательный тракт, нисходящий к двигательным ядрам, проходя по пути через базальные ганглии и РФ ствола. Нужно добавить, что РФ ствола мозга осуществляет модифицирующие влияния - как восходящие (на полушария большого мозга и мозжечка), так и нисходящие (на сенсорные и двигательные ядра). Судя по имеющимся данным, по-видимому, существуют центробежные механизмы, регулирующие сенсорную деятельность организма; соответствующие волокна этого механизма проходят по классическому восходящему сенсорному тракту "против движения". Все упомянутые тракты на самом деле не изолированы, как может показаться из схемы; а взаимодействуют друг с другом. Параллельно с классической последовательностью восходящих нейронов можно заметить и нисходящую систему, связывающую в обратном направлении те же самые узлы промежуточной обработки информации.
В ином виде схема, изображенная на рис. 84, представлена на рис. 85, заимствованном (с некоторыми изменениями) у Брукхарта [34]. На нем схематически представлен мозжечок и его "окрестности" и показано, как развитие новой коры и нового таламуса сопровождалось развитием новых структур мозжечка. О пулах вставочных нейронов спинного мозга (1) уже упоминалось в разд. 5.1. В областях 1 и 2 сосредоточена система управления позой (иногда в этом участвует и зрительная система). Заложить же основы устойчивости позы необходимо, поскольку всякое более сложное целенаправленное поведение отправляется обычно от некоторой устойчивой позы. Экстрапирамидная система (5) является древней (в эволюционном смысле) и богатой перекрестными связями областью и может действовать как сама по себе, так и под управлением коры головного мозга. Возможно, что именно здесь хранятся (врожденные) программы таких двигательных актов, как бег, спаривание, бегство или нападение. Пирамидный путь (5) - система "молодая"; он появляется лишь у млекопитающих, тогда как у птиц сильно развиты другие системы, играющие у млекопитающих менее значительную роль.
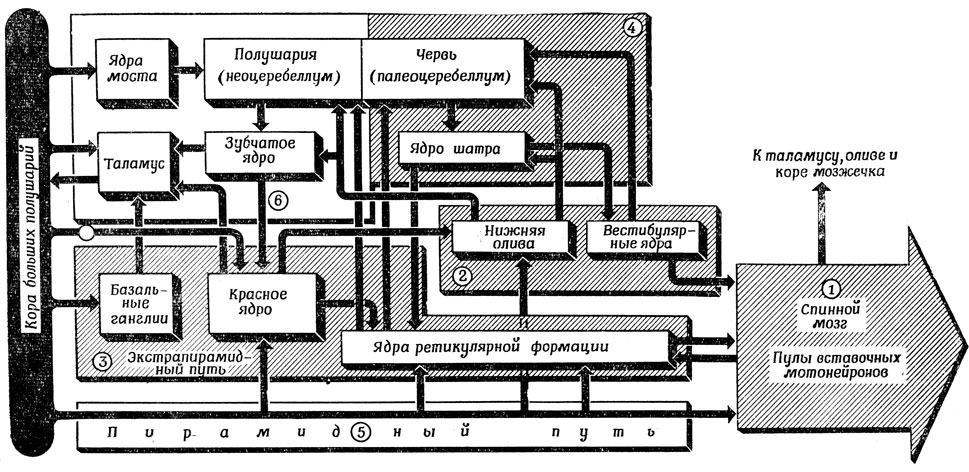
Рис. 85. Схема взаимосвязей между мозжечком и пирамидными и экстрапирамидными путями. Филогенетически более 'древняя' система (блоки 1-4, заштрихованы) дополняется у млекопитающих развитием пирамидного пути (5), таламуса, полушарий мозжечка и связанных с ними структур (6)
Для того чтобы управлять позой (1 и 2) и движением (3 и 5), а также избегать ситуаций, когда пулам вставочных нейронов спинного мозга посылаются противоречивые распоряжения, необходимо какое-то подобие обратной связи между объектом управления и управляющим устройством. Червь мозжечка (палеоцеребеллум 4) получает сигналы из областей 1 и 2, а также от ядер ретикулярной формации и организует канал обратной связи, так сказать, "с черного хода", через ядро шатра. Относительно небольшая часть его выхода уходит через крышу мозжечка в центры управления позой, а также в ретикулярные ядра и пулы вставочных нейронов. Филогенетически более молодые отделы нервной системы используют, по-видимому, новые структуры старых аналогичных отделов для установления связей и организации обратной связи. Например, "новый" мозжечок (6), т. е. два полушария мозжечка, должны координировать ставшие теперь возможными более тонкие изменения позы. Среди входов в эти полушария есть пути от коры головного мозга (проходящие через ядра моста), образующие, по-видимому, часть пирамидного пути, а это наводит на мысль о том, что неоцеребеллум получает информацию о текущих двигательных командах. Однако многие из этих волокон приходят сюда из так называемых ассоциативных двигательных полей, а не только от первичной моторной коры. Выходы "нового" мозжечка проходят через зубчатые ядра. Отметим, что каждое из этих ядер получает такой же входной сигнал, как и соответствующая область коры головного мозга, и обеспечивает выход мозжечка в результате своих собственных расчетов, деталями которых управляет кора. Разница в функционировании областей 4 и 6 состоит в том, что палеоцеребеллум занимается информацией на уровне ретикулярных ядер и ниже, а неоцеребеллум получает более детализированную информацию из базальных ганглиев, гипоталамуса и красных ядер.
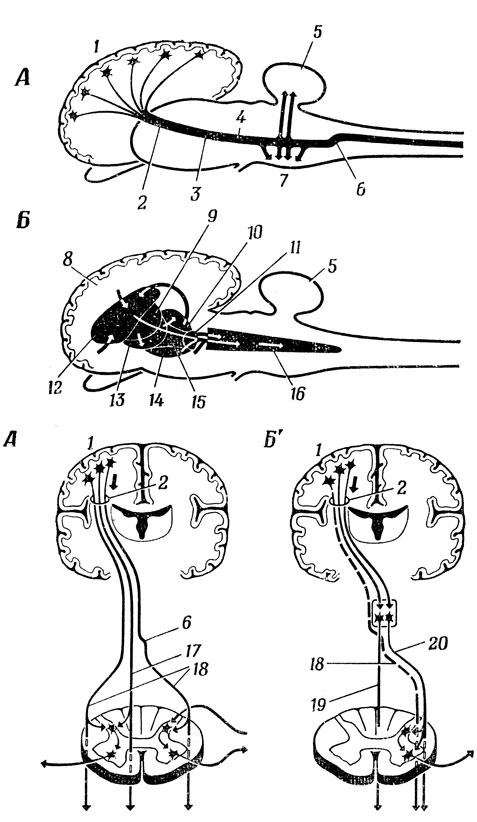
Рис. 86. Схема мозга 'обобщенного' млекопитающего. Показаны те же структуры, которые изображены на рис 85, но пирамидный и экстра- пирамидный пути искусственно разделены. Пирамидный путь - вид сбоку (А) и вид спереди (А'). Экстрапирамидный тракт-вид сбоку (Б) и вид спереди (Б'). 1 - моторная зона коры; 2 - внутренняя капсула; 3 - ножка мозга; 4 - кортико-спинальный тракт; 5 - мозжечок; 6 - перекрест пирамидных путей; 7 - варолиев мост; 8 - кора больших полушарий; 9 - скорлупа; 10 - таламус; 11 - красное ядро; 12 - зубчатое ядро; 13 - бледное ядро; 14 - черная субстанция; 15 - гипоталамус; 16 - покрышка и ретикулярная формация; 17 - передний кортико-спинальный тракт; 18 - латеральный кортико-спинальный тракт; 19 - ретикуло-спинальный и вестибуло-спинальный тракт; 20 - ретикуло-спинальный тракт
Рис. 86-88 дополняют эти схемы, показывая общее взаиморасположение всех упоминавшихся путей. На рис. 86, А представлена нервная система "обобщенного" млекопитающего; мы видим, как аксоны всех клеток сенсо-моторной зоны коры собираются вместе во внутренней капсуле, откуда начинаются волокна кортико-спинального тракта, идущие вниз и отдающие коллатерали в мозжечок при прохождении через варолиев мост. Затем (т. е. ниже) они попадают в перекрест пирамидных путей, где вся масса волокон переходит из одной половины тела в другую (рис. 86, А'). На рис. 86, Б и 86, Б' дана схема экстрапирамидной системы; можно видеть, что главную роль здесь играют подкорковые структуры переднего мозга, а именно базальные ганглии. География базальных ганглиев показана на рис. 87, на котором видно относительное расположение таламуса (блока предварительной переработки сенсорной информации на пути в кору), внутренней капсулы (каналов связи), базальных ганглиев (двигательных систем) и лимбической системы (центры голода, страха, ярости и полового удовлетворения) по мере движения наружу к новой коре. На рис. 87, А слева показан горизонтальный разрез одной половины мозга, а справа - соответствующий вид сбоку. Центральное положение здесь занимает таламус, по обе стороны которого расположены две подсистемы базальных ганглиев: чечевичные ядра и хвостатое ядро. Стрелками показана плоскость сечения, которая расположена достаточно высоко, так что мы не видим зубчика чечевичного ядра на головке хвостатого, но зато нам видна внутренняя капсула как участок белого вещества в форме буквы V, идущий между таламусом и чечевичным ядром и образованный аксонами различных сенсорных путей, а также волокнами различных моторных путей, нисходящими от моторной коры. Другие детали строения лимбической системы показаны на рис. 87, Б и В.
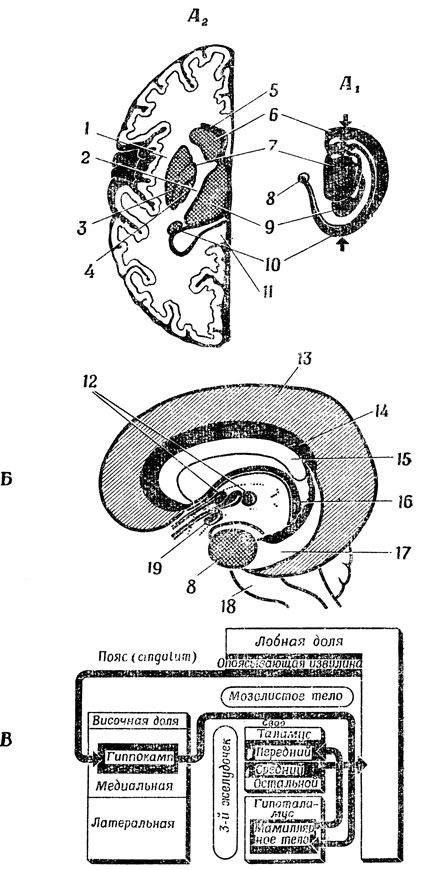
Рис. 87. А. Левая половина мэзга человека: А1 - вид сбоку; А2 - горизонтальный разрез на уровне, показанном на А1 стрелками Б. Лимбическая система обезьяны. В. Схема, объясняющая взаимосвязи между структурами, более реалистично изображенными на А и Б. 1 - наружная капсула; 2 - внутренняя капсула; 3 - скорлупа; 4 - бледное ядро; 5 - мозолистое тело; 6 - головка хвостатого ядра; 7 - чечевичное ядро; 8 - миндалина; 9 - таламус; 10 - хвост хвостатого ядра; 11 - валик мозолистого тела; 12 - ядра таламуса; 13 - новая кора; 14 - поясная извилина; 15 - мозолистое тело; 16 - свод; 17 - гиппокамп; 18 - гипоталамус; 19 - мамиллярные тела
На рис. 88 изображена схема тактильных путей соматосенсорной и сенсо-моторной коры. Здесь мы еще раз убеждаемся в роли таламуса как "промежуточной станции" и видим, что пути, ведущие из таламуса в кору головного мозга, идут также через внутреннюю капсулу (рис. 88, Б). Соматические входные сигналы от головы попадают в таламус через ядро тройничного нерва, а от тела - непосредственно из спинного мозга через латеральный спино-таламический тракт. Наконец, сигналы от проприоцепторов, идущие по задним столбам, попадают в таламус через синапсы ядер нежного и клиновидного пучков. Рис. 88, А еще раз напоминает нам, что ствол мозга связан коллатералями с мозжечком и верхним двухолмием и что в коре сигналы распространяются не только на соматосенсорные, но и на моторные поля и на различные "ассоциативные" области, расположенные в теменной и височной коре. Обратите внимание, что все сенсорные сигналы, за исключением обонятельных, попадают в кору через таламус, где они, по-видимому, подвергаются предварительной переработке, а моторная кора может "разговаривать" со спинным мозгом непосредственно. Однако, кора все же посылает сигналы в таламус, и эти сигналы могут частично "настраивать" сенсорные пути, изменяя передаточные характеристики блоков предварительной переработки информации, расположенных в таламусе. Аналогичным образом существуют возвратные проекции и на двигательные ядра таламуса. На рис. 89 приведена чисто гипотетическая схема формального нейрона, функции которого могут изменяться сигналами настройки, приходящими из коры: вместо обнаружения совпадения нескольких сигналов он просто обнаруживает присутствие любого сигнала.
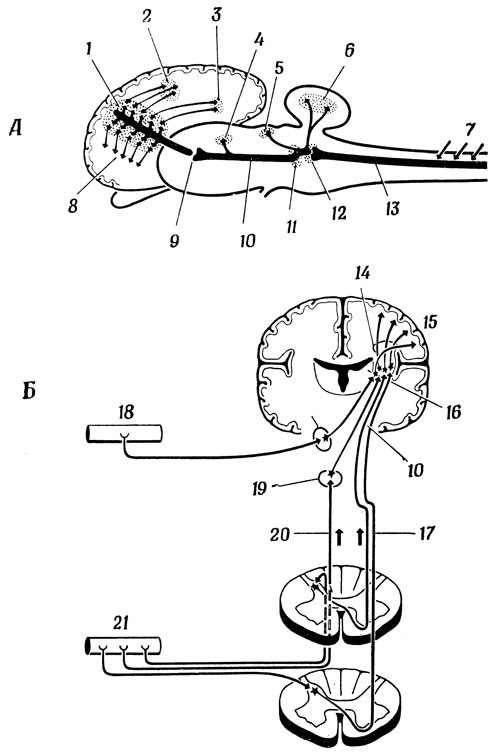
Рис. 88. Две схемы (А и Б) соместетических путей в кору, дополняющие схемы пирамидных и экстрапирамидных систем, показанных на рис. 86. 1 - соместетическая область коры; 2 - ассоциативные волокна теменной доли; 3 - ассоциативные волокна височной доли; 4 - медианное ядро таламуса; 5 - верхнее двухолмие; 6 - волокна к мозжечку; 7 - спинномозговые ганглии; 8 - волокна к моторной области; 9 - вентролатеральное ядро таламуса; 10 - медиальная петля; 11 - перекрест путей в продолговатом мозгу; 12 - ядра нежного и клиновидного пучков; 13 - задний столб; 14 - внутренняя капсула; 15 - постцентральная извилина; 16 - таламус; 17 - передний спино-таламический тракт; 18 - рецепторы головы; 19 - ядра нежного и клиновидного пучков; 20 - задний столб; 21 - рецепторы тела
Заканчивая на этом наше краткое введение в географию мозга, мы хотим обратить внимание читателя на то, что при однодневной экскурсии по большому городу никогда нельзя, успеть познакомиться со всеми его достопримечательностями, да и о том, что удалось увидеть, многое остается неизвестным. Однако, подобно тому как однодневная экскурсия поможет лучше оценить привлекательные черты города при последующих более длительных посещениях, так и наши два раздела, посвященные нейроанатомии, послужат отправной точкой для дальнейших путешествий по научной литературе.
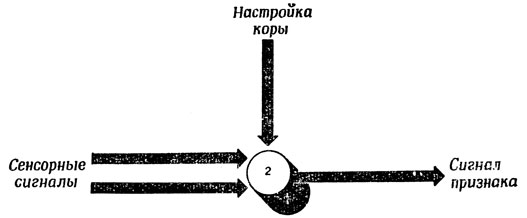
Рис. 89. Гипотетический нейрон, который в зависимости от сигнала настройки может служить как элементом И, так и элементом ИЛИ (ср. с рис. 1)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'