5.5. Распределенное (distributed) двигательное управление
Для того чтобы продвинуть наше понимание того, как теория мозга подходит к проблеме управления движениями, рассмотрим теперь вопрос о том, как распределить по целой пространственно распределенной системе нейронов функции следящей схемы Питтса и Мак-Каллока (воспроизведенной еще раз на рис. 96), которая для каждого φ генерирует преобразование Tφ, приводящее φ к стандартному виду.
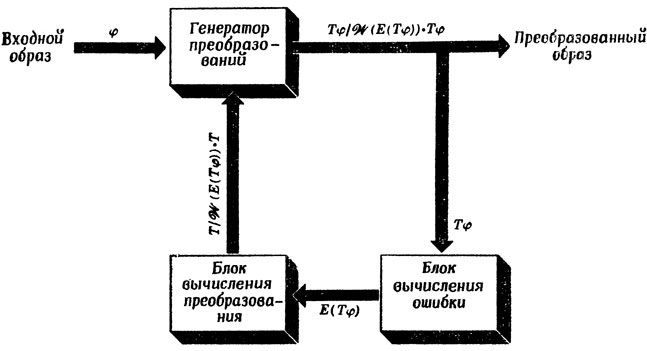
Рис. 96. Обобщенная схема Питтса - Мак-Каллока для преобразования образа к стандартному виду
Как мы показали в разд. 3.3, генератор преобразований не требует памяти: входной образ φ и преобразование T на его входах непосредственно пересчитываются в преобразованный образ Tφ. Не требует памяти и блок вычисления ошибки: в ответ на образ на входе блока на выходе сразу выдается значение ошибки. Генератор же преобразований представляет собой последовательный автомат: если в момент времени t его состоянию соответствовало преобразование T, а на его вход поступил вектор ошибки e, то его новому состоянию в момент времени t+1, а также его выходу будет соответствовать преобразование W(E)•T.
Самое трудное в такой схеме состоит в том, чтобы подобрать подходящую меру ошибки E и соответствующее преобразование W, гарантирующее, что с помощью обратной связи по этой ошибке нам всегда удастся в конце концов привести вход к стандартному виду. Любая теория эволюции и обучения должна рано или поздно столкнуться с задачей формирования таких E и W, которые наилучшим образом отвечают целевой среде системы (мы уже вкратце говорили об этом в разд. 4.2, критикуя ОРЗ).
Для того чтобы связать все это с теорией соматотопически организованных сетей, познакомимся с тем, как Питтс и Мак-Каллок применили свою общую схему к анализу рефлекторной дуги, обеспечивающей такое движение глаз, при котором точка фиксации попадает на центр тяжести распределения яркости зрительного входа. Как известно, эта рефлекторная дуга идет от глаза через верхнее двухолмие к ядрам глазодвигательного нерва, а оттуда в соответствующие мышцы. (Заметим, что на нашем нынешнем уровне знаний относительно системы предварительной переработки зрительной информации в сетчатке мы имеем право заменить слово "яркость" в описании рефлекса словами "общая контурная информация" или "признак".)
Аптер [3, 4] показала, что каждая половина поля зрения кошки (соответствующая назальной половине одного и височной половине другого глаза) топографически проецируется на контралатеральное верхнее двухолмие. Кроме такой "сенсорной" проекции, Аптер изучала и "двигательную", для чего подвергала обработке стрихнином отдельные точки на поверхности двухолмия и давала на сетчатку вспышки света, наблюдая, какую точку зрительного поля фиксировал изменившийся в результате этого взгляд. При этом выяснилось, что "двигательные" и "сенсорные" проекции практически идентичны (см. врезку на рис. 97).
Воспользовавшись этими данными, Питтс и Мак-Каллок построили схему, показанную на рис. 97 и контролирующую направление взгляда. Они обратили внимание на то, что возбуждение некоторой точки левого бугорка соответствует возбуждению, приходящему из правой половины поля зрения, и, следовательно, должно вызывать движение глаза вправо. А в центр взгляд будет направлен тогда, когда возбуждение, приходящее слева, точно уравновесится возбуждением справа. Поэтому Питтс и Мак-Каллок построили свою модель так, что, например, на каждый мотонейрон, управляющий мышечными волокнами левой внутренней и правой наружной прямых мышц, сокращение которых вызывает движение соответственно левого и правого глазного яблока вправо, поступает суммарное возбуждение клеток, расположенных в тонкой поперечной полоске, в составе левого бугорка. Это обеспечивает необходимое возбуждение правой наружной и левой внутренней прямых мышц, а соответствующие мышцы поворачивают глаза вправо. Реципрокное торможение через коллатерали аксонов от ядер мышц-, антагонистов, возбуждаемых аналогичным образом клетками другого бугорка, создает суммарное возбуждение, вычитаемое из первого. Вычисление вертикальной координаты подобного квазицентра тяжести осуществляется аналогичным образом., Конечно, такие вычисления могут выполняться и при участии волокон комиссур, соединяющих аналогичные точки контралатеральных тектальных отделов, а не ядер глазодвигательного нерва. Движение глаз в такой схеме прекращается тогда и только тогда, когда точка фиксации совпадает с центром тяжести.
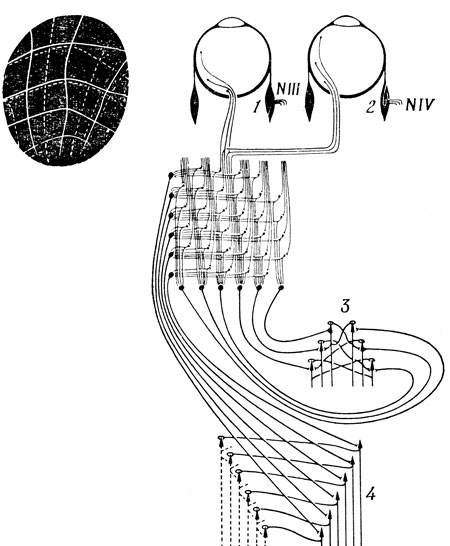
Рис. 97. Схема Питтса - Мак-Каллока для объяснения рефлекторного контроля движений глаз. Глаз остается неподвижным только тогда, когда активность в обоих бугорках верхнего двухолмия сбалансирована. 1 - внутренняя прямая мышца левого глаза; 2 - наружная прямая мышца правого глаза, 3 - мотонейроны, управляющие вертикальным движением; 4 - мотонейроны (N III, N VI), управляющие левой внутренней и правой наружной прямыми мышцами глаза. Слева вверху показана сенсорная (пунктирная линия) и моторная (сплошная линия) проекции Аптер на правой бугорок верхнего двухолмия кошки
Необходимо подчеркнуть, что рефлекс, для которого мы только что построили весьма грубую, но полезную модель, при нормальном функционировании управляется еще и "высшими центрами". Например, выбор участка зрительного поля, на который направлен взгляд, может диктоваться "интересом", а рефлекс при этом определяет лишь выбор точки фиксации в данном участке (ср. с системой автоподстройки в радиоприемнике); взгляд может оставаться фиксированным на некоторой точке до тех пор, пока она не будет "достаточно" изучена. Напротив, неожиданная яркая вспышка может подавить все остальные компоненты операции усреднения и на короткое время взять верх над рефлекторным контролем, вынуждая организм оказать хотя бы мимолетное внимание новой ситуации.
Хотелось бы отметить, что даже если включить в уравнения, описывающие схему Питтса - Мак-Каллока, демпфирующие члены, препятствующие возникновению незатухающих колебаний глазных яблок относительно их положения равновесия, у этой схемы все равно останется один серьезный недостаток: она по самой своей сути следящая, тогда как рефлекторная "переброска" взгляда в направлении вспышки света существенно баллистическая. На самом деле движения глаз человека могут управляться как по баллистической, так и по следящей схемам. Обычно человек, рассматривающий некоторую сцену, сначала фиксирует взгляд на одной точке зрительного поля, а затем его глаза совершают саккадические (термин для баллистического движения глаз) движения, фиксируя взглядом другую точку, и так до тех пор, пока не будет просмотрено достаточное число точек для того, чтобы составить себе представление о внешней обстановке. Однако в других ситуациях, например наблюдая за проезжающим мимо автомобилем, прежде чем перейти улицу, человек фиксирует взгляд на этом объекте и следит за ним. У приматов различные области коры могут модулировать активность верхнего двухолмия, и Биззи удалось выяснить, что в одной из них, в так называемом фронтальном зрительном поле, находящемся в лобной доле коры, имеются три типа клеток: клетки типа I работают при баллистических движениях глаз, типа II - при следящем режиме этого движения, а остальные клетки занимаются не столько движениями глаз, сколько движениями головы. Возможно, что при более тщательном исследовании подобная ситуация будет обнаружена и в верхнем двухолмии. В любом случае создается впечатление, что схема Питтса - Мак-Каллока больше подходит для описания следящего, чем баллистического режима.
Чтобы исправить это, перейдем к изучению другой модели, предложенной Брайтенбергом и Онесто [31], которая описывает баллистические движения, управляемые вычислительной машиной с распределенными функциями. (Следует сказать, что Брайтенберг и Онесто придумали свою модель для описания работы мозжечка. Однако впоследствии было получено так много новых данных о мозжечке, что сохранить эту модель для коры мозжечка без коренной переработки оказалось невозможным. Критику различных моделей мозжечка можно найти в работе Бойлза и Арбиба [30], а здесь вряд ли следует вдаваться во все подробности. Я не стремлюсь к тому, чтобы создать модель функционирования той или иной подсистемы мозга. Скорее мне хотелось бы показать некий плодотворный способ моделирования функций мозга. Руководствуясь этим, я рассказываю здесь о моделях, содержащих новые принципы организации, надеясь стимулировать дальнейшие исследования по выяснению биологических механизмов, воплощающих эти принципы в нервной системе, или возможностей их использования в конструкции систем управления для роботов.)
При стрельбе основную роль играют две силы: взрывная сила заряда, толкающая снаряд к цели, и сила торможения, возникающая в момент, когда снаряд попадает в цель (если бы цель отошла в сторону, снаряд не смог бы остановиться в той точке, в которую целились). Баллистические движения животных также основаны на подобном "релейном" принципе управления. Опыты с быстрым сгибанием и разгибанием конечности в суставах показали, что по времени активация мышц составляет лишь небольшую часть движения и что продолжительность таких периодов активации, по-видимому, не зависит от продолжительности движения. Вначале возникает короткая вспышка активности, во время которой мышца-агонист активно сокращается, а мышца-антагонист расслабляется, и в результате соответствующая конечность приобретает ускорение; затем наступает период покоя, после чего происходит новая вспышка активности, вызывающая сокращение мышцы-антагониста и торможение движения.
Эксперименты Биззи с движениями глаз выявили систему, в которой в "пусковой момент" возникает возбуждение мышцы-агониста при полной пассивности мышцы-антагониста, и так продолжается вплоть до "финального момента", когда обе мышцы возвращаются на фоновый уровень возбуждения, соответствующего новой схеме. Создается впечатление, будто одновременно включается вспышка активности агониста и сигнал управления следящей системой, выводимой на новый уровень, причем первый из видов активности преобладает над вторым вплоть до его отключения. Во всяком случае, и в той, и в другой ситуации продолжительность движения определяется, по- видимому, в основном временем подачи сигнала "стоп" относительно сигнала "пуск", которое диктуется мозгом, а не внешней средой, как при полете снаряда. Таким образом, Брайтенберг и Онесто предложили схему преобразования пространства во время, поскольку в ней положение входного сигнала (в котором закодировано положение искомой цели) определяет момент выхода (проявляющийся в том, что мы "ударяем по тормозам"). В их схеме (рис. 98) имеется линейная цепочка выходных клеток, выходные каналы которых организованы таким образом, что возбуждение любой из этих клеток вызывает возбуждение мышцы-антагониста и тормозит баллистическое движение. Входные волокна этих клеток распадаются на две группы, образующие две одинаковые линейные цепочки, причем их расположение на прямой соответствует различным значениям суставного угла. Каждое волокно первой группы - С-волокна - соединяется с единственной выходной клеткой. Волокна же второй группы - М-волокна - ветвятся так, что каждое из них оказывается соединенным со всеми выходными клетками. Скорость распространения возбуждения по этим параллельным волокнам такова, что время, необходимое на передачу возбуждения от одной выходной клетки к другой, равно времени, затрачиваемому суставом на соответствующее изменение суставного угла, предполагая постоянство угловой скорости движения в этом суставе.
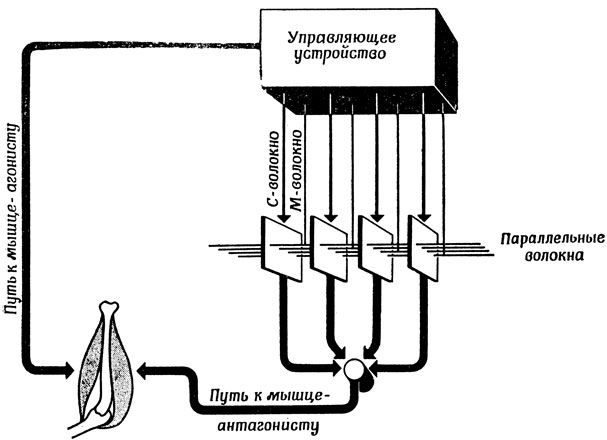
Рис. 98. Схема Брайтенберга - Онесто для управления баллистическим движением. В ней пространственное разделение возбужденных С-волокон и М-волокон преобразуется в разделение возбуждения мышц - агониста и антагониста во времени
Регулятор описываемой системы запускает баллистическое движение, генерируя три сигнала. Если говорить лишь о движениях в суставе, то первый из этих сигналов возбуждает мышцу - агониста, что и приводит к началу движения. Другой сигнал по С-волокну поступает на выходную клетку, соответствующую исходному положению сустава, а третий - по М-волокну на выходную клетку желаемого положения. (Читателю предлагается самому придумать необходимую систему связей для управления саккадическими движениями глаз.) Если предположить, что выходная клетка может реагировать на сигнал, пришедший по параллельным волокнам, только при условии, что она получила до этого сигнал по С-волокну, то понятно, что из всех выходных клеток может возбудиться только та, которая соответствует возбужденному С-волокну, а время ее возбуждения будет соответствовать ее расстоянию до клетки, возбужденной М-волокном. Такая схема вызовет торможение движения за счет возбуждения мышцы-антагониста как раз в нужный момент времени.
Заметьте, что в этой схеме мы освобождаем регулятор от необходимости "знать", в каком положении находится сустав, организовав контур обратной связи таким образом, что он осуществляет непрерывный контроль за этим состоянием и обеспечивает возбуждение именно того С-волокна, какое нужно.
Конечно, мы не утверждаем, что предложенная здесь модель точно соответствует тому, как на самом деле нервная система управляет движениями. Мы лишь продемонстрировали некую правдоподобную схему (рис. 99), возможно соответствующую организации одной из подсистем нервной системы позвоночных; на этой схеме выбор места ввода входного сигнала на поверхности управляющих элементов может кодировать задание для мышц. Более того, можно ожидать, что (по аналогии с результатами объединения схем Питтса - Мак-Каллока и Брайтенберга - Онесто) при возбуждении целого множества точек на такой поверхности система перейдет в состояние, соответствующее "центру тяжести" этого множества.
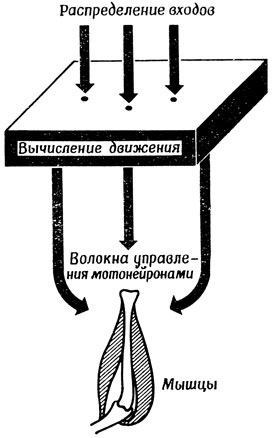
Рис. 99. Схема распределенной системы управления движением, реализующей механизмы типа показанных на рис. 97 и 98. Пространственно закодированное положение цели преобразуется в соответствующую последовательность команд, а весь набор входов вызывает движение к 'центру' закодированных целей
Необходимо отметить, что, доведя эту схему до ее логического конца, мы пришли бы к иерархической системе. Например, при фиксации новой точки в пространстве увеличение угла отклонения от среднего положения потребует сначала соответствующего движения одних только глаз, затем головы и глаз вместе, а затем глаз, головы и всего туловища. Поэтому выходной сигнал вычислительного устройства, управляющего такими движениями, должен управлять не каким-то одним суставом, а целой иерархией регуляторов более низкого уровня, поведение которых, естественно, в свою очередь должно зависеть и от регуляторов позы низшего уровня, о которых говорилось в предыдущем разделе. Заметим, что эта схема должна также контролировать скорость движения и т. п. В качестве первого грубого приближения можно предположить, что подобный подход выработался в процессе эволюции в результате совершенствования "схем", управляющих следящими движениями изнутри, а не с помощью сенсорных каналов.
Как мы подробнее выясним в разд. 6.3, поведение любого организма зависит главным образом от того, в какой мере он способен установить правильные взаимоотношения с теми частями внешней среды, с которыми он собирается взаимодействовать. В разд 4.4 мы уже видели, что при движении организм должен корректировать свою внутреннюю модель, чтобы она могла служить прочной основой для планирования его будущих взаимодействий со средой. Чтобы связать эти соображения со структурами, показанными на рис. 96, для которых целью или "стандартной формой" служит "объект, расположенный в месте, удобном для взаимодействия", рассмотрим животное, в зрительное поле которого попал какой-то объект, а остальное зрительное поле затянуто туманом. Тогда две различные ситуации:
1) животное приблизилось к объекту на 1 м,
2) объект приблизился к животному на 1 м
вызовут одинаковую стимуляцию сетчатки. Но несмотря на такое совпадение стимуляции, во "враждебной" среде (а именно в такой среде протекала наша собственная эволюция) лишь в ситуации 2 объект важен для животного и требует внимания, ориентации в его сторону и идентификации источника возбуждения. Точно так же, если объект "не отстает" от животного, оно должно обратить на него внимание, хотя у организма и нет зрительной обратной связи, сообщающей ему о его собственном движении или о движении объекта. Это подводит нас к одному важному понятию: хотя воспринимающая система получает информацию в виде изменений стимуляции, изменения, вызванные собственными движениями животного, необходимо "исключить" из числа наблюдаемых изменений; в противном случае они могли бы замаскировать важные перемены во внешней среде.
Изящное экспериментальное доказательство существования такого "исключения" можно найти в работе фон Хольста и Миттельштедта [173]. Оно подтверждает гипотезу, согласно которой восприятие зависит не только от сенсорных входов как таковых, но в не меньшей степени от разницы между изменениями, произведенными собственными движениями организма, и изменениями на его рецепторных поверхностях. Фон Хольст и Миттельштедт проводили свои эскперименты на мухах, но после описания их исследовании мы приведем и подтверждающие данные, относящиеся к человеку.
Если ограничить подвижность мухи так, чтобы она могла лишь поворачиваться относительно некоторой неподвижной точки, а затем поместить ее во вращающийся барабан, на стенках которого нарисованы вертикальные полоски, позволяющие отчетливо видеть вращение (рис. 100), то муха будет поворачиваться в сторону вращения, как бы "отслеживая" его. Если затем остановить барабан и поместить по одну сторону от мухи источник соблазнительного для нее запаха, то она повернется к этому источнику. Однако при этом ее повороте происходит такое же смещение полосок относительно сетчатки ее глаз, как если бы барабан вращали в обратном направлении. Тем не менее муха не изменяет в связи с этим направления своего вращения, как следовало бы ожидать, если бы мы предположили, Это ее реакция на сенсорную стимуляцию не зависит от ее собственного движения. Для такой способности мухи "игнорировать" вращение в противоположном направлении можно предложить по крайней мере два удовлетворительных объяснения. Одно из них состоит в том, что во время собственного движения муха просто "блокирует" все зрительные входные сигналы, а, согласно другому объяснению, она действительно "вычитает" свое собственное движение из наблюдаемого движения внешней среды. Фон Хольст и Миттельштедт изящно исключили возможность первой гипотезы, повернув голову мухи относительно ее туловища на 180°, что можно сделать, не нарушая нервных связей между глазами и телом. Такая муха, раз начав вращаться, продолжает делать это до тех пор, пока не падает от изнеможения; это наводит на мысль, что вместо того, чтобы "блокировать" зрительные входы, она действительно вычитает планируемое движение из наблюдаемого (которое теперь из-за неестественного положения головы оказывается строго противоположным планируемому) и реагирует на образующееся несоответствие так, как если бы это и было истинным движением внешней среды.
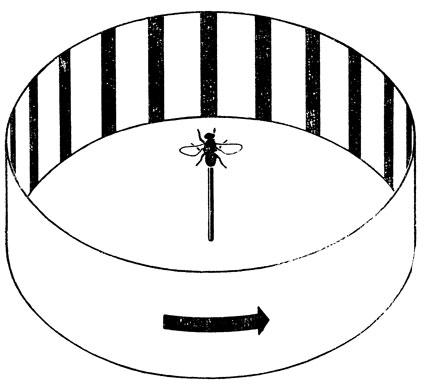
Рис. 100. Экспериментальная установка, которая демонстрирует, что муха чувствует разницу между собственным движением и изменениями, вызванными движением вовне
Данные о человеке подтверждают этот вывод. Когда человек впервые надевает инвертирующие призмы, при каждом движении головы окружающий его мир начинает "прыгать" в глазах. И здесь, как и в предыдущем случае, инверсия приводит к изменению направления возбуждения сетчатки; оно становится противоположным тому, которое обычно связывают с этим движением. Занятно, однако, что человек может в конце концов адаптироваться к такому обращению, а муха не может. Читателю предлагается предсказать, что ощущает человек с недавно возникшим параличом глазных мышц, когда он "хочет" перевести взгляд с одного предмета на другой.
Познакомившись с экспериментальными подтверждениями способности животных в восприятии какого-либо одного параметра учитывать эффект своих собственных движений, мы должны заметить, что у млекопитающих с их сложно регулируемыми рецепторами, осуществляющими многомерную связь со столь же сложными эффекторами, возникают еще более тонкие проблемы. Мы считаем, что им необходимы механизмы, построенные по тому же принципу, что и схема на рис. 101.
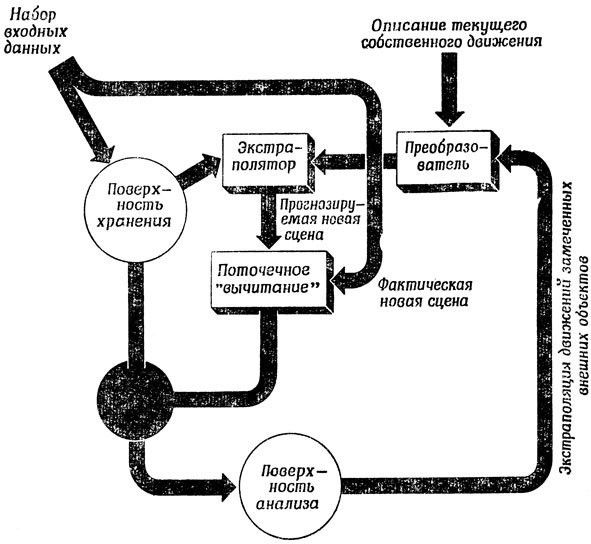
Рис. 101. Схема, позволяющая игнорировать несущественные изменения в сенсорном входе. На поверхность анализа поступают в качестве входов только те аспекты набора входных данных, воспринимаемых системой в целом, которые не были предусмотрены в результате анализа своих собственных движений и движений тех объектов во внешней среде, которые система уже заметила
На рис. 101 входной набор данных готов поступить на "поверхность хранения", где он вскоре заменит предыдущий подобный вход. Новый входной набор поступает одновременно в механизм, где происходит сравнение некоторых признаков этой структуры с соответствующими признаками "прогнозируемой новой сцены". Этот прогноз составляется путем коррекции предыдущей входной сцены (записанной на "поверхности хранения"), с учетом как собственных движений системы в текущий период, так и экстраполяции движений опознанных объектов внешнего мира. Таким образом, те участки новой сцены, в которых она отличается от старой только по причине собственного движения системы и продолжения движения попавших в эти участки объектов, согласуются с соответствующими участками "прогнозируемой новой сцены", что в свою очередь вызывает подавление соответствующих отрезков путей, ведущих от "поверхности хранения" в первую "поверхность анализа". На самом деле здесь, по-видимому, используется смешанная баллистико-следящая стратегия. "Баллистическое" движение, вызванное преобразователем, может быть вполне точным, и прежде чем пытаться устанавливать соответствие и принимать решение о торможении некоторых связей, может потребоваться определенная тонкая настройка. Необходимость такого уточнения была впервые замечена Мак-Кеем, который впоследствии опубликовал обзор своих работ [157].) Связи, идущие от тех участков новой сцены, которые не соответствуют прогнозу, не тормозятся на пути к "поверхности анализа", а именно на этих участках система должна сосредоточить свое внимание. Надо подчеркнуть, что схема на рис. 101 - не более чем предварительный набросок и что блок "преобразования" и экстраполятор - это устройства с высокой степенью распараллеливания функций, способные по-разному преобразовывать различные части набора входных данных. Более того, мне совершенно не ясно, сколь важную роль в подобных процессах играет прямое эфферентное управление на первых этапах входных путей, и в какой мере сами выходные комплексы "поверхности анализа" через посредство обратной связи влияют на работу блока "преобразования" и экстраполятора.
Для Питтса и Мак-Каллока в их модели двухолмия самым главным было то, что она служила примером реализации предложенной ими схемы, согласующимся с нейрофизиологическими данными. А нам хочется сделать главный упор на то, что эта модель демонстрирует возможность существования соматотопически организованных сетей, в которых нет "руководящего нейрона", предписывающего поведение системы в целом, а вместо этого динамические системы эффекторов с помощью связывающих их взаимодействий нейронов извлекают выходную траекторию из популяции нейронов, каждый из которых располагает лишь локальной информацией о том, как должна вести себя система. И главная мысль настоящей книги состоит в том, что центральным элементом теории мозга должно стать изучение именно таких сетей. Развитию этого подхода и посвящена следующая глава.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'