5.4. Иерархическая структура управления двигательной системой
В предыдущих разделах этой главы мы выяснили, что системы связи спинного мозга играют центральную роль в формировании движений конечностей, и познакомились с проблемами координации этих движений, компенсации нерегулярностей внешних условий и обеспечения коррекций позы, позволяющих животному сохранять равновесие при сложной двигательной активности. Однако в этом разделе, вместо того чтобы сконцентрировать внимание на детальном разборе реальных схем, мы хотим показать, что для ряда задач стратегическое планирование на достаточно высоком уровне не должно задавать детали выполнения этого плана на мышечном уровне.
Если мы хотим написать какое-то слово один раз карандашом на клочке бумаги, а другой раз - на стене длинной малярной кистью, которую приходится держать двумя руками, то очевидно, что в каждом случае используются совершенно разные группы мышц. Так чем же объяснить, что в обоих случаях проявляются характерные особенности нашего почерка, даже если кистью мы пишем впервые в жизни? Здесь, пожалуй, полезно вспомнить об интерпретирующих системах для вычислительных машин (разд. 3.2). Найдя целесообразным составлять программы не на машинном языке, т. е. не в терминах реальных операций, выполняемых схемами вычислительной машины, а на некотором языке более высокого уровня, мы можем снабдить машину управляющей программой, называемой интерпретирующей системой и позволяющей переводить каждую команду программы высокого уровня на язык машинных программ и выполнять их. Имея это в виду, мы воспользуемся нашей метафорой "человек - это машина" и представим себе, что, обучаясь в детстве письму, мы научились не только производить ряд нужных движений, чтобы водить карандашом, зажатым в руке, но и сформировали себе представление о механизме письма на языке более высокого уровня - в терминах пространственно-временной последовательности штрихов. Продолжая эту мысль (а читатель должен иметь в виду, что все эти соображения носят чисто гипотетический характер), мы можем предположить, что в. мозгу накапливаются программы высокого уровня в терминах последовательностей, направленных в разные стороны движений и относительных положений, не уточняющие ни абсолютного положения, в котором должно осуществляться это движение, ни тех частей тела, которые его реализуют. Одновременно можно выдвинуть гипотезу, что в процессе обучения формируется внутри нас и интерпретирующая система для подобных программ высокого уровня, а точнее - по одной интерпретирующей системе на каждую из систем управления нашего тела. Поэтому при наличии той или иной программы высокого уровня любая такая система может осуществлять весьма сложные движения, хотя она никогда прежде их не выполняла. Это объяснение, по-видимому, достаточно хорошо подходит для нашего примера с надписью на стене. Конечно, многие вопросы остаются при этом открытыми; какие именно механизмы в мозгу вырабатывают программу высокого уровня из навыка, приобретенного только одной системой, и строят интерпретирующие системы для разных систем? В какой мере оправданно вводить столь четкое разграничение между языком высокого уровня, интерпретирующей системой и машинным языком самих эффекторных систем?
Во всяком случае, предыдущие рассуждения позволяют рассматривать сложную двигательную активность как результат "срастания" в единое целое нескольких субструктур, каждая из которых имеет дело лишь с ограниченным числом аспектов общей проблемы. В связи с этим одной из важных задач теории мозга становится вычленение таких автономных субструктур моторного поведения и анализ способов их сочетания, приводящего к координированным действиям. Этой проблемой достаточно глубоко занимался Грини [86-89]. Чтобы в полной мере представить себе суть этой проблемы синтеза, мы можем, следуя Грини [88], рассмотреть зрительный дисплей, сконструированный Дертоузосом [56, 57]; этот дисплей дает возможность генерировать картинки, управляя лучом катодно-лучевой трубки. Очевидно, самый общий подход состоит в том, чтобы описывать картинку на экране, задавая яркость экрана в каждой его точке. Таким образом можно построить любое изображение. Однако в большинстве "интересных" случаев более эффективным оказывается другой подход, основанный на конструировании необходимого изображения из стандартных кривых, каждая из которых зависит к тому же от нескольких параметров. С некоторым подобием такого "синтетического" подхода мы уже познакомились в разд. 4.1, говоря о "кассетной" метафоре для оперативной памяти.
Дисплей Дертоузоса основан на втором подходе (рис. 92). Если сигнал на входе 2 равен нулю, то при сигнале "Рисовать" на входе 1 на экране дисплея луч вычерчивает диагональ, Регулируя значения четырех параметров с помощью входа 2, мы получаем возможность в ответ на сигнал "Рисовать" на входе 1 вычертить на экране "практически любую" гладкую кривую (это напоминает универсальность лекала для чертежника). Следующий шаг состоит в том, чтобы создать подсистему, генерирующую необходимые сигналы настройки для параметризованного семейства примерно десяти стандартных кривых. Входами такого регулятора более высокого уровня будут названия соответствующих кривых, их ориентация, время появления на экране, начальная точка, размер и т. п. Подобным способом мы можем построить иерархическую систему на высшем уровне, которой не придется заниматься нудной и кропотливой работой по вычислению сигналов, подаваемых на отклоняющую систему, для каждого момента времени.
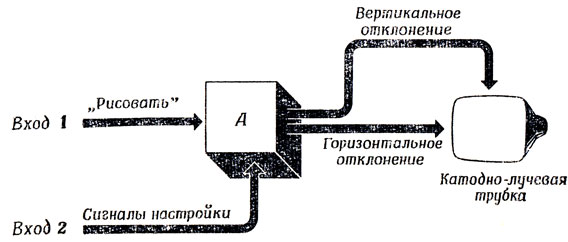
Рис. 92. Зрительный дисплей Дертоузоса. Объяснения в тексте
При таком подходе мы не отказываемся ни от одной идеи обычной теории управления. В локальных регуляторах мы должны пользоваться обратной, а возможно, и упреждающей (т. е. использующей модель для предсказания того, что должно случиться; ср. разд. 3.4) связями. И хотя точный размер выигрыша нельзя оценить количественно, пока мы не разберемся лучше в теории сложности вычислений, цель наша состоит в том, чтобы сократить объем вычислений на высшем уровне за счет загрузки более локальных, менее универсальных, менее автономных и более простых регуляторов.
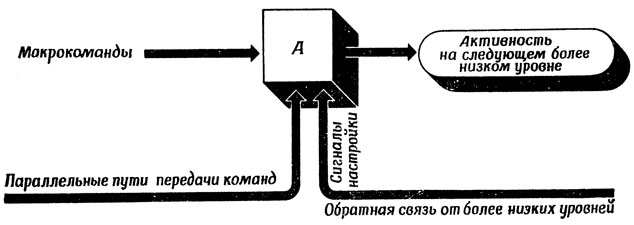
Рис. 93. Обобщение схемы, изображенной на рис. 92. Тонкая настройка движения, вызванного макрокомандами, достигается при помощи как сигналов обратной связи от низших уровней моторной системы животного, так и параллельных каналов
Возвращаясь к вопросу о двигательной системе животных, модифицируем рис. 92 в виде рис. 93. Здесь на любом иерархическом уровне предпринимаемые действия запускаются соответствующей макрокомандой и настраиваются в основном сигналами о местных условиях, но, возможно, также еще и сигналами с ближайшего верхнего уровня. От макрокоманды требуется только, чтобы она давала действию "правильное направление", т. е. обеспечивала приближенную траекторию движения, которую можно было бы скорректировать, не выходя за пределы возможностей сигналов настройки. Хорошим примером такого положения вещей может служить понятие эталонной позы, относительно которой выполняются соответствующие движения. Контур обратной связи с помощью спинальных рефлексов может помешать животному упасть, несмотря на изменение условий опоры. Поэтому команды о начале движения не должны нести информации о том, что произойдет с центром тяжести после того, как будет поднята одна из ног. Сигналы "настройки" позы вызовут (если это будет необходимо) такое смещение центра тяжести, которое обеспечит сохранение устойчивости. Уточнение (необходимое, например, для управления движениями пальцев при игре на рояле) такой стабилизированной с помощью обратной связи макротраектории может осуществляться по нескольким параллельным каналам. (Конечно, в этом случае макрокоманды должны обеспечивать траекторию, близкую к устойчивой. Например, если команды высшего уровня заставляют животное перенести три ноги за край пропасти, трудно ожидать, что сигналы настройки помогут восстановить устойчивую стабильную ситуацию.)
Полезно отметить, что многие типы поведения, которые достаточно трудно моделировать на вычислительной машине, могут оказаться простейшими действиями нервной системы и тела. Наличие силы тяжести дает нам возможность опустить руку вниз, не производя никаких расчетов. Кроме того, на требования, предъявляемые к расчетам в нейронных сетях, оказывает влияние зависимость между формой конечностей и их функциями. Анатомы и особенно эволюционисты много работают над тем, чтобы выяснить корреляции между изменениями структуры скелета и изменениями функций организма. Очевидно, что есть много разных способов сборки суставов, в результате которой получится система, способная осуществлять определенные действия, но для некоторых из таких структур управлять этими действиями будет очень просто, а для других - весьма сложно. В теории автоматов доказаны теоремы о том, как изменение кодирования входных сигналов может привести к резкому изменению времени, необходимого автомату на вычисления. Для каждой функции существует, по-видимому, оптимальный способ кодирования информации, при котором система вычисляет эту функцию быстрее всего (ср. с нашими рассуждениями о предварительной обработке зрительных образов в разд. 2.4). Аналогично структурные свойства мускулатуры и скелета в значительной степени предопределяют объем вычислений, необходимых для того, чтобы гарантировать плавную работу суставов и простоту взаимодействия организма с окружающим его внешним миром. В связи с этим особое значение для нас может приобрести такой "старомодный" раздел механики, как кинематика и теория машин и механизмов: добавляя кинематические соображения к информационным, мы можем научиться строить из брусков и стержней, соединяемых шарнирами, такие структуры, которые могли бы вычерчивать желаемую траекторию, воспроизводящую без чрезмерно сложных расчетов последовательные фазы предписанного движения. Таким образом, хотя наше внимание практически целиком сосредоточено на распараллеливании процессов переработки информации по всей активной сети, с тем чтобы обеспечить управление действием, комбинируя текущие входные сигналы с прошлым опытом, необходимо отдавать себе отчет и в том, что решаемые вычислительные задачи зависят не только от характера предварительной переработки входных сигналов, но и от структуры тех исполнительных "механизмов" на выходе, которые реализуют соответствующее действие на практике.
Завершим наши теоретические рассуждения кратким перечнем преимуществ и недостатков иерархической организации системы управления, в которой наибольшая эффективность высших уровней достигается за счет снижения сложности производимых на этих уровнях вычислений (ср. с разд. 4.1, где рассматривались преимущества и недостатки внутренних моделей). К преимуществам следует отнести: 1) увеличение времени на решение творческих задач (планирование), связанных с появлением новых ситуаций; 2) экономичность использования имеющегося объема памяти, связанную с отсутствием необходимости запоминать все мелкие детали; 3) увеличение скорости и простоты обучения эффективному поведению в новых (или аналогичных) ситуациях (если только они не слишком непривычны). Наличие локальных систем управления позволяет очень быстро организовать несложные специфические действия, вроде отдергивания руки от горячей печки. Еще одно преимущество заключается в том, что по сравнению со структурой с единственным уровнем управления в иерархической структуре переток информации гораздо меньше.
Но у иерархической системы есть и свои недостатки. Так, если бы нас интересовала лишь эффективность чисто моторной части движения (без учета вычислительной работы в высших центрах), то можно было бы найти лучшую структуру и лучшую последовательность возбуждения мышц при условии, что наши расчеты учитывали бы все подробности ситуации. И как уже можно было понять из примера с попыткой одновременно хлопать себя по голове и гладить по животу, иерархическая структура системы управления коренным образом сужает репертуар возможных поведений животного. Это означает, что некоторые необычные изменения внешней обстановки могут поставить животное в условия, с которыми оно не справится. Более того, обучаться "необычным" задачам (например, новым "па" современных танцев) оказывается весьма сложно. Однако совершенно ясно, что как для животного, так и для робота большинство встречающихся задач допускает декомпозицию на знакомые подзадачи, а эта постепенная (уровень за уровнем) отработка поведения обычно гораздо более эффективна, чем любая попытка заменить команды высшего уровня некоторой последовательностью команд на машинном языке.
Занимаясь исследованиями эволюции нейронных систем, мы испытываем сильное искушение предположить, что на рис. 92 изображена система, способная извлечь все преимущества из иерархической организации управления, при которой различные контуры управления с обратной связью в спинном мозгу и выше обеспечивают тонкое управление позой, а схема на рис. 93 показывает, как устроен дополнительный механизм, позволяющий преодолеть один из недостатков иерархических систем, отмеченный выше, и открывающий возможность "прямого вмешательства сверху" как дополнительный источник тонкой подстройку основанный на использовании опыта и результатов обучения.
Опыт, позволяющий выявить это различие и связать его с материалом о системах с обратной связью в спинном мозгу (разд. 5.1), был поставлен Навасом и Старком [180]. Однако, не вдаваясь в детали этого опыта, расскажем, что можно обнаружить, если исследовать ситуацию, изображенную на рис. 94. Возвращаясь к рис. 80, следовало бы ожидать, что в момент падения груза в протянутую руку мышечные веретена растянутся и это приведет к усилению возбуждения α-мотонейронов, в результате чего угол в локтевом суставе практически вернется к прежнему значению. На самом же деле время реакции оказывается дольше, чем нужно для сгибательного рефлекса, и по крайней мере на первой фазе компенсаторного движения мышечные веретена в нем, по-видимому, не участвуют. Создается впечатление, что мы имеем дело с примером прекрасно отработанного двигательного акта: осознав наличие значительного возмущения, мы используем баллистическое движение, направленное на его компенсацию, а мышечные веретена включаются лишь в самом конце для тонкой регулировки конечного участка траектории и для восстановления системы тонкой компенсации незначительных возмущений.
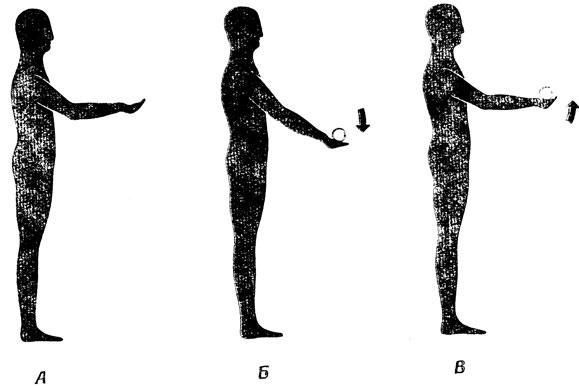
Рис. 94. Как нервная система могла бы реагировать на неожиданное изменение нагрузки. Рука находится в требуемом положенин (А), затем под действием груза она опускается вниз (Б), после чего вычисляется сигнал баллистического управления, вызывающий обратное движение руки к требуемому положению, причем окончательная его 'доводка' (В) обеспечивается следящей системой
Завершим этот раздел некоторыми данными о специфичности нейронных связей (прекрасный обзор по этому вопросу принадлежит Джекобсону [127]), которые должны еще раз напомнить нам, что "подстройка" двигательного поведения действительно состоит из двух составляющих: 1) "локальной подстройки", осуществляемой с помощью контуров, проходящих по спинному мозгу и стволу мозга, и 2) "изменения стратегии" и "добавления мелких деталей", для реализации которых требуется, по-видимому, образование новых разделов мозга, подобно новой коре млекопитающих. Эксперименты по нервной специфичности показывают, что, хотя в "высших центрах" мозга и возможно обучение, "низшие центры" ствола и спинного мозга имеют генетически предопределенную структуру, обеспечивающую основу для нормальной функции; если в процессе регенерации поврежденных нервов эта структура нарушается, то восстановление ее нормальной функции возможно лишь при условии образования другой, пусть измененной, но "удачной" структуры, функция которой может компенсироваться под влиянием высших центров и при условии существования соответствующих высших центров.
Для того чтобы оценить опыты Вейсса [252], показывающие, что нервные связи не есть результат обучения - и что они могут не измениться, даже если становятся неадаптивными, - полезно вернуться к рис. 37, где изображена весьма упрощенная схема управления ногой с четырьмя мышцами (рис. 36); возбуждение единственного on-нейрона "высшими центрами" запускает ритмическое движение, которое мы назовем "двигательным отсчетом (*)". Для наших нынешних целей нам требуется отметить одну важнейшую особенность любой схемы, способной воспроизвести "двигательный отсчет (*): если мотонейрон Ad подключить к мышце Ab, а мотонейрон Ab - к мышце Ad, то активация схемы вызовет "двигательный отсчет" в обратном направлении.
Рассмотрим ситуацию, изображенную на рис. 95, где передние ноги тритона поменяли местами, так что они обращены назад. Тритон обладает достаточной способностью к регенерации, поэтому пересаженные конечности приживляются и в них прорастают спинномозговые нервы. Можно было бы ожидать, что возникающие при этом взаимосвязи между нервами и мышцами окажутся совершенно случайными и животное либо вовсе не сможет управлять своими передними конечностями, либо после соответствующей "сортировки связей" научится в конце концов пользоваться ими, хотя и несколько неуклюже из-за их необычного расположения. Однако на практике получается обратное: животное начинает выполнять передними конечностями движения, которые совершенно не соответствуют их новому расположению, отвращая его от пищи и, напротив, подталкивая по направлению к неприятным стимулам. Вейс [252] по этому поводу писал: "Подлинное движение в обратную сторону происходит только тогда, когда другие органы передвижения, например хвост и задние конечности, парализованы или удалены. При наличии задних конечностей суммарный эффект сводится к непрерывному противоборству между передними и задними конечностями, причем первые толкают тело назад, а вторые - вперед с одинаковой силой. В результате, животное раскачивается взад и вперед, не будучи в состоянии сдвинуться с места. Животные были совершенно беспомощны, и хотя некоторые из них жили более года, т. е. в течение еще довольна долгого времени после метаморфозы, их поведение ничуть не изменялось".

Рис. 95. Неприятные последствия пересадки правой передней конечности на место левой и наоборот для движения тритона
Учитывая наши рассуждения о нейронных схемах, реализующих "двигательный отсчет (*)", эти результаты позволяют предположить, что в опыте Вейсса отводящая мышца оказалась соединенной с мотонейронным пулом приводящей мышцы и наоборот, хотя изменение роли этих мышц после того, как правую и левую конечности поменяли местами, требовало переключения нервных связей. (Такой анализ может показаться слишком прямолинейным, но пока нам этого вполне достаточно.)
Возможно, что читатель давно уже задает себе недоуменный вопрос: "Но разве тритон не мог заметить, что он движется не в ту сторону, и принять необходимые меры?" Поскольку эксперимент отвечает на этот вопрос отрицательно, полезно попытаться найти источник высказанного недоверия. В самом деле, если воспользоваться простейшим примером и предположить, что человек едет на машине, у которой обычное положение передачи соответствует заднему ходу и наоборот, то, вероятнее всего, ему в конце концов надоест удаляться от цели и он обучится переключать передачу по-новому. При этом он не только поймет, что всякий раз, когда хочется двигаться вперед, нужно переключить передачу на обратную, но и должен будет: 1) осознать, что прямая и обратная передачи соответствуют строго противоположным направлениям движения; 2) догадаться, что прямой передаче соответствует движение назад; 3) выдвинуть и проверить гипотезу о том, что для движения вперед нужно включить обратную передачу, и, наконец, после того как эта гипотеза подтвердится, 4) запомнить это и 5) суметь использовать вытекающие выводы. И даже принимая во внимание, что задачи 1 и 3 можно отбросить, поскольку на использование обратной передачи можно натолкнуться случайно в процессе проб и ошибок (что, впрочем, требует, чтобы время от времени мы отходили от установленных процедур), все же остается совершенно ясным, что действие, казавшееся простым проявлением здравого смысла, на самом деле предполагает последовательное решение ряда задач, а у нас нет никаких априорных оснований предполагать, что мозг любого животного пригоден для их решения.
В качестве последней и весьма яркой иллюстрации необоснованности подобных ожиданий предположим, что в вычислительной машине перепутались коды команд сложения и вычитания. И даже если мы станем бить по машине ногой всякий раз, как она ошибается, это не поможет ей переписать программу. Нам придется признать, что для того, чтобы такие воздействия приводили к цели, необходимо снабдить машину специальными датчиками и предусмотреть, чтобы наше негодование учитывалось во время вычислений, но - что еще более важно - нам придется ввести в машину весьма сложную управляющую программу (и надеяться, что перестановка кодов не вывела ее из строя), которая в ответ на удар ногой начнет испытывать различные способы коррекции программ нижнего уровня до тех пор, пока удары не прекратятся.
Опыты Вейсса показывают, что у тритона просто нет такого механизма высокого уровня. Его эволюция происходила вереде, где нет биолога-экспериментатора, и поэтому для того, чтобы выжить, он не нуждается в способности к столь сложным процессам адаптации. Тритон - животное с раз и навсегда "запаянной" программой: поставьте перед ним пищу, и его мозг "скомандует" спинному мозгу "вперед", но если такая команда вызывает движение в обратном направлении, приспособиться к этому он уже не может. Подобные наблюдения заставляют скорректировать наивные представления о поведении как реакции на стимулы (тесно связанные, между прочим, с теорией случайных соединений нейронов), согласно которой любое поведение формируется в результате обучения тому, что на часто повторяющийся стимул нужно отвечать определенной реакцией. Необходимо ясно отдавать себе отчет в том, что многое в нашем поведении "запаяно" (а соотношение между тем, что "запаяно", и тем, что является результатом обучения, зависит от "уровня" животного), а также в том, что плюс к обучению реакциям на стандартные стимулы обучение на других уровнях организовано гораздо сложнее; поэтому, когда человек пишет эссе, его генетическое наследие дает ему лишь субстрат, рефлексы помогают водить ручкой по бумаге или стучать пальцами по клавишам пишущей машинки, а собственно содержание его произведения возникает лишь на самых верхних уровнях организации его мыслительных процессов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'