6.1. Принципы организации
Для того чтобы понять, как "работает" нервная система, необходимо иметь правдоподобные представления о том, как она устроена. Конечно, мы в состоянии регистрировать активность отдельных клеток, но если бы мы предприняли измерение активности почти всех клеток мозга, это дало бы нам, с одной стороны, слишком много, а с другой - слишком мало. Слишком много потому, что мы потонули бы в этой невообразимой массе данных, а слишком мало - потому что мы не смогли бы разглядеть в них важных закономерностей. Поэтому в этом разделе мы постараемся сформулировать некоторые "принципы" организации, которые могли бы помочь нам преодолеть это препятствие и сделать задачу экспериментатора по сбору данных более обозримой.
В начале шестидесятых годов Мак-Каллок собирался написать "Трактат по философии нейрофизиологии". Он написал к нему введение, которое мы здесь воспроизводим, чтобы показать, какое огромное влияние его взгляды на нейрофизиологию оказали на нашу теорию мозга. К сожалению, остальные главы книги Мак-Каллока так и остались ненаписанными. Однако многие из его идей читатель сможет найти в труде "Материализация мозга", в котором собраны работы Мак-Каллока.
Введение в трактат по философии нейрофизиологии
0.0. Человек начал разбираться в своем поведении, в своей психологии за две тысячи лет до того, как он перестал считать, что мозг существует лишь затем, чтобы охлаждать кровь, и за четыре тысячи лет до того, как выяснил, что мозг - это не студенистое вещество, из которого в результате конденсации образуются нервные волокна. Понимание устройства мозга может оказаться полезным для изучения поведения человека, поскольку оно объясняет нам самого человека, но оно не в состоянии учесть то, на что направлено это поведение. Поэтому нейрофизиология не достигает уровня психологии, когда речь идет об отношениях между тремя объектами, например следующего типа: "A ведет себя так, как если бы B казалось ему C". Мы не собираемся предлагать никакого исчисления таких триадических отношений и вообще какой бы то ни было логики психологии.
0.10. Нам хотелось бы лишь предложить логику, пригодную для нейрофизиологии как науки о нервной системе. Нервная система имеется только у многоклеточных животных и образована из клеток, называемых нейронами. Гистологически нейроны обнаруживаются благодаря своему сродству к некоторым красителям. Морфологически они образуют единую систему, единую сеть, поскольку в этой системе каждый нейрон связан со всеми остальными. Электрически нейроны отличаются тем, что они проводят электрические импульсы. Нейроны образуют единую систему и с физиологической точки зрения, поскольку появление импульсов в одном нейроне влияет на аналогичные процессы в нейронах, с которыми он связан, и зависит в свою очередь от появления импульсов в других соседних нейронах. Эта сеть устроена таким образом, что на каждый эфферентный периферический нейрон (т. е. каждый нейрон, управляющий эффектором, будь то мышца или железа) может косвенно, по разным путям, проходящим через сеть, влиять любой периферический афферентный нейрон (т. е. каждый нейрон, на который непосредственно действует какой-то рецептор, будь то сенсорная клетка или какой-то другой датчик). При этом через каждый нейрон проходит по крайней мере один такой путь.
0.11. Место соединения нейронов - синапс, где один из них оказывает влияние на другой, - функционирует в основном только в одном направлении. Поэтому распространение влияний по любому пути происходит лишь в этом направлении, Однако в сети имеется также много контуров, т. е. замкнутых путей, и даже в периферических афферентных нейронах характер возбуждения и передачи импульсов обычно зависит от поведения других нейронов. Более того, сами рецепторы испытывают влияние эффекторов, управляемых эфферентными периферическими нейронами. От контуров, замыкающихся через рецепторы и поддерживающих в них возбуждение, зависит наша жизнь: благодаря им мы дышим. Аналогичные контуры в недрах системы обеспечивают контакт с внешним миром: благодаря им мы воспринимаем то, что происходит вокруг. Те контуры, выходной сигнал которых одновременно поступает и на вход с обратным знаком, т. е. наши рефлексы, и другие отрицательные обратные связи в мозгу, обеспечивают стабильность системы, возвращая ее к тому состоянию, из которого ее вывело возмущение. Возвращение к исходному состоянию - вот функция каждого такого контура, его цель. Установленное таким образом состояние определяет финал и конечную цель работы такого контура, его энтелехию.
0.12. Наши рецепторы чрезвычайно чувствительны к малейшим изменениям интенсивности специфических форм энергии или концентраций некоторых веществ. Поэтому такие изменения обычно вызывают последовательность импульсов в примыкающих афферентных периферических нейронах, состояния которых определяются нервной системой в целом в зависимости от того, где находится нейрон и о каком времени идет речь. Таким образом, последовательности импульсов в любом и каждом афферентном периферическом нейроне сообщают нервной системе о происходящих вокруг событиях. Поэтому генерируемые последовательности импульсов играют для нервной системы роль полезных сигналов при условии адекватности внешних стимулов и состояния воспринимающих эти стимулы нейронов. Последовательности импульсов, вызванные любыми другими причинами, составляют то, что инженеры называют шумами. Полезные сигналы и шумы, которые непременно должны различать как инженеры, так и нейрофизиологи, описываются (но не различаются) физиками на языке изменения материи и энергии. Они различаются функционально - в зависимости от того, помогают или не помогают нервной системе решить стоящие перед ней задачи.
0.13. Поскольку происходящие в окружающем нас мире события довольно однородны, а наши рецепторы расположены в довольно тесной близости друг к другу, практически любой адекватный стимул воздействует одновременно на целый комплекс однотипных датчиков. А так как каждый датчик может влиять на несколько афферентных периферических нейронов, испытывая в свою очередь влияние нескольких других датчиков из данного комплекса, сигналы, генерируемые многими из этих нейронов, сильно коррелируют между собой. Именно благодаря такой избыточности мы и распознаем объекты восприятия и такие их свойства, как "голубой", "сладкий", "твердый" и "пронзительный".
0.132. Любой внутренне связанный источник адекватной стимуляции, движущийся относительно какой-то поверхности, например сетчатки или кожи, последовательно воздействует на соседние рецепторы. Это вызывает возбуждение соседних, перекрывающихся групп афферентных периферических нейронов, у которых вызванные этим стимулом изменения последовательности импульсов продолжаются дольше, чем вызвавший их эффект. Благодаря подобной корреляции одновременных сигналов мы воспринимаем конкретные объекты, характеризующиеся наличием траектории, например летящую искру, оцарапавшую нас веточку или гудок промчавшегося мимо поезда. На любой афферентный периферический нейрон соматической сенсорной системы или любую биполярную клетку сетчатки и воздействуют датчики более чем одного типа, хотя воздействие, оказываемое на последовательность импульсов в этом нейроне, может меняться в зависимости от типа датчика. Поэтому изменения свойств движущегося объекта, если только они не слишком велики или не слишком неожиданны, не мешают нам идентифицировать его на основании его траектории.
0.133. Если бы не отмеченная выше избыточность, трудно было бы представить себе, каким образом животному удается находить своих родителей, добывать пищу, обходить препятствия, защищаться от врагов, находить себе брачного партнера или заботиться о своем потомстве. Она позволяет нам идентифицировать и классифицировать объекты, несмотря на то что мы воспринимаем их на фоне несущественных адекватных стимулов из внешнего мира и шумов, вызванных несовершенством работы датчиков. Эти вопросы нас здесь не интересуют, но, решая все эти проблемы, природа наградила нас нервной системой с сильно коррелированными входами, и поэтому мы должны рассматривать сигналы в афферентных периферических нейронах как результат кодирования с высокой избыточностью. И забывать об этом нельзя.
0.141. Более того, центральные окончания этих нейронов таковы, что они обеспечивают эффективную проекцию окрестности рецепторов в окрестность центральных нейронов-реципиентов и соответствующие окрестности пересекаются между собой. Центральное расположение наших эфферентных периферических нейронов также образует аналогичную топологию эффекторов, и возникающие при этом отображения тоже избыточны.
0.142. Простейшая рефлекторная дуга образуется рецептором растяжения, воздействующим па эфферентный периферический нейрон растянутой мышцы и вызывающим ее сокращение. Но это не единственный контур распространения возбуждения. Другой контур, проходящий через те же афферентные нейроны, подключает в работу нейроны, управляющие мышцами-антагонистами для растянутой мышцы в одном или нескольких суставах, и вызывает в них уменьшение напряжения. Все другие последовательные дуги из большего числа последовательно соединенных нейронов содержат некоторые промежуточные связи, которые часто называют вставочными нейронами.
0.143. По аналогичному принципу устроены и дистантные рецепторы, расположенные в голове. Так, фоторецепторы топологически проецируются на биполярные клетки, а те в свою очередь - на ганглиозные клетки глаза. Сигналы этих клеток идут в верхние бугорки четверохолмия и через него в кору головного мозга. Они попадают также и в латеральное коленчатое тело и через него снова в кору головного мозга. Все эти пути топологически связаны один с другим. И если бы не такая многоканальность связи на всем протяжении пути от рецепторов к эффекторам, нам пришлось бы передавать с помощью какой-то последовательности сигналов и информацию о расположении места восприятия, с тем чтобы именно в это место была направлена соответствующая реакция. А это - "локальный адрес" Лотце* - составляет в случае зрения 20 бит информации на каждую из миллиона ганглиозных клеток.
* (См., например, R. H. Lotze "Medizinische Psychologie oder Physiologie der Seele", Leipzig, 1852.)
0.145. Многоканальность связи позволяет сохранить этот локальный адрес всюду, кроме тех частей мозга, в которых сигналы из многих или даже всех участков организма собираются воедино: либо для того, чтобы принять решение, касающееся поведения всего организма в целом (как, например, в ретикулярной формации), либо для того, чтобы распознать объект, воспринимаемый с помощью нескольких сенсорных модальностей, либо, наконец, для того, чтобы выработать навыки или суждения, зависящие от такого целостного восприятия, как это имеет место в коре головного мозга.
Опираясь на эти идеи Мак-Каллока и материал первых пяти глав этой книги, мы попробуем вкратце сформулировать несколько принципов, которыми будем руководствоваться при попытке построить теорию мозга.
I. Мы пытались, в частности в разд. 2.1 и 4.1, разобраться в основных функциональных свойствах, которыми, по-видимому, должна обладать любая система, способная приспособиться к сложной внешней среде, будь то робот или животное. В этом смысле мы, возможно, идем дальше Мак-Каллока и не отрекаемся, как он это делает в пункте 0.10, от всяких попыток построить логику, пригодную не только для нейрофизиологии. Пытаясь понять, почему животным удается целенаправленно и эффективно передвигаться в постоянно меняющемся внешнем мире, важно отметить, что изменения этого внешнего мира все же не выходят за известные пределы. Поэтому животному (или роботу) нет необходимости уметь пользоваться своими эффекторами в каждой из необозримого множества возможных комбинаций. Оно может решать многие задачи на подсознательном, полурефлекторном уровне, пользуясь хорошо отработанным репертуаром взаимодействий. Быть может, понятие внутренней модели даст нам возможность подступиться к отношениям типа "А ведет себя так, как если бы В казалось ему С", о которых говорит Мак-Каллок в пункте 0.0, хотя, конечно, мы ни в коей мере не готовы предложить общую логику триадических отношений.
По сути дела, наше понимание функций организма основано на представлении, согласно которому восприятие всегда нацелено на действие. При этом предполагается, что система, которая собирается взаимодействовать с переменчивой окружающей средой новым и адаптивным образом, должна построить и постоянно корректировать свою внутреннюю модель взаимодействий с миром; поэтому восприятие в значительной мере зависит от этой модели и направлено на ее совершенствование - а тем самым и на совершенствование будущих действий - в той же степени, в какой оно определяет текущее действие.
Изучая последствия такой направленности восприятия на действие и на построение моделей, мы выяснили, что среди всех соотношений между организмом и окружающим миром особую роль должны играть пространственные отношения. Мы обнаружили также, что восприятие именно в силу того, что оно открывает системе различные возможности действия, одновременно и создает для него дополнительные трудности, связанные с необходимостью устранения избыточности потенциальных команд, или, иначе говоря, с необходимостью выбрать не более одного из целого множества доступных организму действий.
II. Мы выделили несколько аспектов, по которым мозг существенно отличается от машин, созданных человеком. Один из таких аспектов связан с элементной базой: мы спрашиваем, что следует из того, что мозг представляет собой сеть нейронов. Мы добавляем к этому, что мозг - это живая система и вместе с остальными частями организма развился из одного яйца. Многие свойства мозга легче понять, исходя из требований биологического развития, а не из функции взрослой особи. Другое отличие имеет исторический характер: мы не можем не делать различия между мозгом человека, возникшего из мозга других животных в процессе эволюции, одним из последних достижений которой было развитие речи, и управляющими устройствами робота, который даже в простейшем случае умеет в полном смысле слова манипулировать символьной информацией.
Слишком часто вследствие переоценки роли языка мы считаем, что любое наше взаимодействие с внешним миром требует языкового опосредования приблизительно следующего вида:
1) увидев объект, назови его,
2) по названию объекта назови подходящее действие,
3) действуй согласно этому названию.
Эта простейшая схема усложняется, однако, учетом таких соображений, как необходимость взвешивания альтернатив. Но так или иначе такой подход приводит к структурам типа показанной на рис. 102, А. Многие специалисты по искусственному интеллекту идут именно по такому пути, изыскивая алгоритмы, рассчитанные на обычные "последовательные" ЭВМ и основанные на преобразованиях цепочек символов, а не пытаются понять механизмы мозга, лежащие в основе человеческого разума.
Центральная (и, как мне кажется, ошибочная) идея, заложенная в схеме 102, А, состоит в том, что все входные сигналы можно свести к различным сочетаниям нескольких удачно выбранных слов, которые можно преобразовывать с помощью некоторого централизованного руководящего органа в команды, управляющие нашими мышцами. И хотя, добавив к этому несколько подходящих контуров обратной связи, можно получить модели, позволяющие анализировать многие аспекты человеческого поведения, нам хотелось бы подчеркнуть, что существуют весьма совершенные стратегии обеспечения сенсомоторных отношений, не требующие такого суперрегулятора; поэтому может оказаться весьма полезным рассматривать вербальное опосредование лишь как еще один уровень иерархической структуры (рис. 102, Б). Здесь вербальное опосредование может непосредственно управлять выходом системы, но во многих аспектах разумного поведения, не носящих явно языкового характера, без него можно вполне обойтись. Это достигается либо путем передачи управления низшим центрам, либо путем перестройки лежащих в основе такого поведения сенсомоторных механизмов, которые, вообще говоря, при вычислении действий не пользуются в явном виде вербальными кодами.
В этой книге мы обычно игнорируем "лингвистический уровень" и вместо этого опираемся на мысль, что "смысл" входа "для организма определяется теми взаимодействиями, в которые он может вступать с объектом, представляемым этим входом, а эти действия зависят не только от того, что это за объект, но и от того, где он находится.
Система, представленная на рис. 102, Б, анархична в том смысле, что в ней нет голову или центра, который единовластно руководил бы ее вычислениями. Это распределенная вычислительная машина с высокой степенью распараллеливания действий. В связи с этим необходимо понять, как можно производить вычисления в сложной сети динамически взаимодействующих подсистем с высокой степенью распараллеливания. Поняв это, нам будет легче оценить, как различные повреждения мозга могут отразиться на поведении. В 1929 г. Лешли [146] писал, что снижение эффективности поведения крыс в лабиринте, вызванное удалением различных участков коры, по-видимому, не зависит от того, какой именно участок был удален, а определяется лишь количеством удаленной коры. Это позволило ему сформулировать два "закона": закон массы, согласно которому снижение эффективности зависит от массы удаленного участка, и закон эквипотенциальности, утверждающий, что каждый участок мозга может вносить одинаковый вклад в процесс решения задач. Все эти данные казались несовместимыми со взглядом на мозг как на вычислительную схему со строго распределенными функциями в раз и навсегда сформированной структуре. Однако их, быть^может, удастся совместить, представив, себе вычислительную схему, основанную на координации многих подпрограмм, работающих одновременно и параллельно. Ведь нередко для вычислений требуется лишь часть подпрограмм. И хотя в общем случае исключение одной из подпрограмм должно понизить эффективность системы, для некоторых задач недостающая подпрограмма может быть несущественна, а значит, ее устранение может лишь спасти систему от ненужных затрат времени на ее отработку в момент, когда нужно заниматься другими делами. Р. Уайт (личное сообщение) повторил опыты Лешли, но вместо того, чтобы оценивать снижение эффективности поведения крыс по одному-единственному параметру, он пытался выяснить источник такой деградации. Одна крыса может деградировать из-за того, что у нее появляется тенденция все время поворачивать влево, другая легко отвлекается, третья может сидеть неподвижно почти все время, но без всякого труда находит путь через лабиринт всякий раз, когда она "соизволит" попробовать. Таким образом, закон эквипотенциальности действует лишь тогда, когда мы оцениваем изменения в поведении довольно грубо; в самом деле, удаление различных подсистем может вносить совершенно различный вклад в нормальный уровень поведения.
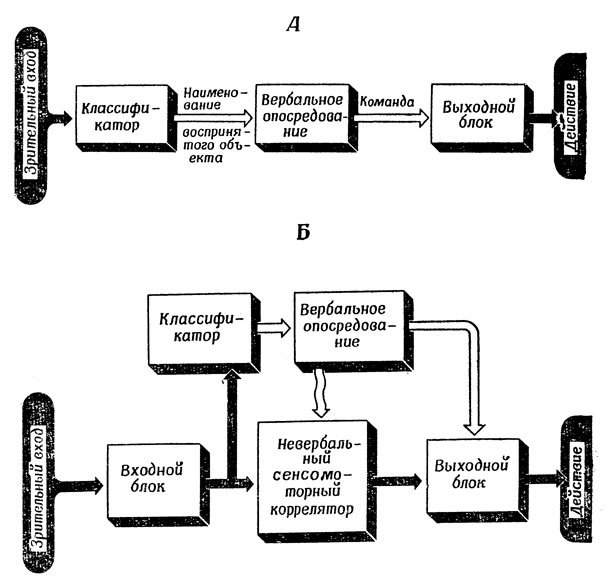
Рис. 102. Ошибочный подход к восприятию и действиям, основанный на вербальном опосредовании (А), которому мы противопоставляем более реалистическую схему (Б). Хотя по схеме Б вербальное опосредование и может оказывать прямое влияние на выход, в большинстве случаев это не обязательно для разумного поведения, поскольку эту задачу берут на себя низшие центры или же ее выполнение обеспечивается просто путем перестройки сенсомоторного аппарата
III. Поэтому стоит заняться исследованием основной архитектоники таких систем, что, по нашему мнению, может оказаться полезным для поиска методов компактного описания вычислений, распределенных по сети из 10 миллиардов нейронов. Основное свойство такой структуры отмечено Мак-Каллоком в пункте 0.143, где он пишет, что в мозгу имеется много путей, каждый из которых топологически связан с другими. Мы будем формулировать это, говоря, что "мозг есть многослойная соматотопическая вычислительная машина", и посвятим остаток данного раздела обсуждению этого принципа; при этом следует отметить, что в соответствии с пунктом 0.145 Мак-Каллока этот принцип, как бы полезен он ни был, применим не во всех случаях.
В разд. 3.3 мы высказали предположение, что, хотя нам, может быть, и удалось бы сконструировать робот, "мозг" которого был бы организован как централизованный преобразователь (θdθ)-θ в соответствии с рис. 43, целесообразность централизованного процессора начинает казаться сомнительной, если представить себе, что его выход нужно "разыгрывать" на целой системе мотонейронов, как это имеет место в биологических системах. Красота схемы Питтса - Мак-Каллока для верхних бугорков четверохолмия (разд. 5.5) заключена в той простоте, с которой она демонстрирует (по крайней мере в рассматривавшемся случае), что без централизованного процессора вполне можно обойтись и что все вычисления можно осуществить распределенным образом на одном или нескольких слоях, разделяющих структуры входных и выходных элементов.
Как мы уже знаем из разд. 2.4, Леттвин, Матурана, Мак-Каллок и Питтс [150] выяснили, что большинство ганглиозных клеток сетчатки лягушки можно отнести к одному из четырех классов, например к классу "детекторов границы" или к классу "детекторов движущегося закругленного края". Здесь же нам хотелось бы подчеркнуть, каким образом информация, поступающая из детекторов четырех типов, распределяется в области тектума. Окончания этих детекторов образуют четыре разных слоя, расположенных один над другим и обладающих следующими свойствами: 1) различным слоям соответствуют детекторы разных типов, 2) в каждом слое сохраняются пространственные соотношения между исходными клетками (т. е. определенное направление по слою соответствует определенному движению относительно сетчатки), 3) окончания, лежащие в этих четырех слоях друг над другом, относятся к ганглиозным клеткам, рецептивные поля которых перекрываются. Это еще один яркий пример нейронной специфичности, которая создает структурную основу для функций мозга. Следует отметить, что в подобных взаимосвязях между двумя слоями могут соблюдаться грубые пространственные соотношения (вверх и вниз, но не поперек), но нарушаются соотношения размеров. Например, в одном слое человеческого мозга, получающем информацию от рецепторов прикосновения, рецепторам пальца может отводиться больше места, чем рецепторам всего туловища, поскольку для того, чтобы управлять тонкими манипуляциями, мозгу необходима подробная сенсорная информация из пальцев. Такая взаимосвязь между двумя слоями клеток называется соматотопической (от греч. сома - тело и топос - место), так как она сохраняет информацию об участке тела, где возникло возбуждение, по мере движения от рецепторов в центральную нервную систему. Чем дальше мы отходим от периферии, тем менее выраженной становится такая взаимосвязь, но мы по-прежнему можем основывать свои исследования на предположении, что в двух соседних слоях такие соотношения сохраняются. Этим мы хотим сказать, что один из полезных способов структурного описания кажущегося хаоса многих частей мозга состоит в том, чтобы описывать эти части как совокупность взаимосвязанных слоев, в которых положение на слое является основным индикатором функционального содержания активности данной клетки, а анализ одной полоски такого слоя может обеспечить понимание функции слоя в целом.
Рассматривая соматотопические связи между слоями в таком распределенном вычислительном устройстве, читатель должен иметь в виду, что мы употребляем здесь термин "соматотопический" в чрезвычайно широком смысле. Для входных путей зрительной системы расположение на слое служит колом расположения в зрительном поле относительно глаза, но (и для нас это имеет принципиальное значение) эта кодировка соотнесена к системе координат нашего тела как результат сформированных в процессе индивидуального развития связей между сетчаткой и мозгом. В слуховой системе положением на слое кодируется частота стимуляции вблизи периферии, в тактильной системе - положение на теле; в последнем случае термин "сомалотопическая связь" приобретает свой буквальный смысл. И хотя термины "ретинотопическая связь" или "тонотопическая связь" точнее бы соответствовали другим ситуациям на периферии, нам кажется, что по мере движения в центральную нервную систему кодирование должно отражать соотношения между организмом и внешним миром в более общей форме. Что же касается выходных путей, то положение в слое кодирует там, возможно, расположение цели какого-либо движения. Можно ожидать, что по мере перехода от периферии к слоям мозга, весьма удаленным от преобладающего влияния какой-то одной сенсорной модальности или какого-то одного вида действий, положение в слое будет весьма мало зависеть от расположения в теле. Однако мы полагаем, что и при этом положение в слое все еще кодирует какие-то основные параметры функции клетки. И именно в этом, возможно чрезмерно широком, смысле позиционного кода мы будем говорить о соматотопии даже для весьма глубоких слоев, далеких от периферии. Более того, в мозгу имеются структуры, и, возможно, ретикулярная формация одна из них, где представления о многослойности неприменимы.
Мы не собираемся фанатически отстаивать справедливость утверждения: "Мозг есть соматотопически организованное многослойное вычислительное устройство". Мы просто хотим воспользоваться им как удобным девизом, напоминающим нам о том, что настало время в полную силу использовать в наших теориях мозга понятие соматотопичности, уже давно играющее важную роль в анатомии и физиологии. Даже в тех структурах, которые не являются слоистыми, положение нейронов может играть роль, которой нельзя пренебречь при моделировании его вклада в общую функцию данной структуры.
Именно это мы имели в виду, когда в конце гл. 1 выражали надежду, что нам удастся убедить читателя в жизнеспособности исследований, направленных на создание моделей "распределенных деенаправленных вычислений в многослойных сома- тотопически организованных машинах".
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'