6.2. Комплексы выходных признаков
Ранее (особенно в гл. 5) мы уже отмечали, что спинной мозг организован таким образом, что обычные наборы стимулов, которые он получает из высших центров (например, из ствола мозга и мотосенсорной коры), вызывают не подергивания отдельных мышц, а координированное движение, в которое может быть вовлечено несколько мышц, как станет ясным, если попытаться пошевелить только одним средним пальцем ноги. Сенсорные признаки, о которых уже говорилось в разд. 2.4, дополняют двигательные признаки, экономно описывающие объекты в терминах наших возможных взаимодействий с ними. Мы отмечали также (например, говоря о вычислительном устройстве для управления движением в разд. 5.5), что каждая мышца состоит из целой популяции мышечных волокон; поэтому в крупномасштабных движениях (в отличие, например, от тонких движений пальцев у человека) участвует целая популяция мотонейронов. Тогда в соответствии с нашими представлениями о деенаправленности восприятия мы можем предположить, что распознавание текущей обстановки предполагает такое возбуждение коры (это утверждение остается пока спекулятивным), которое, если бы оно передалось спинномозговым центрам, привело бы к целому ряду попыток взаимодействия организма с окружающей средой, причем каждая из этих попыток более или менее соответствовала бы данной среде. Поэтому одна из центральных задач "синтеза" нервной системы - устранение избыточности потенциальных команд, обеспечивающее в случае взаимной несовместимости конкурентных структур поведения проявление в каждый момент времени лишь одной из них и притом той, которая больше всего подходит к создавшейся для организма ситуации. В гл. 7 мы познакомимся с двумя возможными механизмами устранения такой избыточности.
Мы договорились выше рассматривать многие уровни мозга как многослойное вычислительное устройство, в котором положение на слое определяет пространственное расположение источника стимуляции, способного возбудить клетки в этом участке слоя. Предположим поэтому, что и в "выходных структурах" мозга положение на слое определяет, в какой точке пространства произошло бы взаимодействие, если бы спинальные центры могли "реализовать" поведение, управляемое данным участком. Заметим, что, поскольку одно и то же движение может быть вызвано сенсорной стимуляцией многих различных типов, у некоторых клеток "двигательных слоев" могут быть широчайшие "рецептивные поля", если классифицировать их по типу входов, способных воздействовать на их активность. Поэтому мы различаем (рис. 103*) : 1) "сенсорные слои", в которых состояние внешней среды закодировано в виде совокупности определенных признаков и координат той части пространства, к которой они относятся, а дешифровка информации осуществляется с помощью нескольких слоев предварительной обработки (см. разд. 2.4) картины возбуждения рецепторов под влиянием внешних стимулов и 2) "двигательные слои", в которых состояние внешней среды закодировано в виде совокупности определенных "выходных признаков" или "двигательных подпрограмм" и координат той части пространства, в которой они подвергнутся преобразованию, а дешифровка информации и ее преобразование в координированные движения мышц происходят путем их обработки в одном-двух слоях нейронов мотосенсорной коры и спинного мозга. Что касается областей, лежащих между "сенсорными" и "двигательными слоями", то очевидно, что необходимость многосторонней интеграции данных в процессе планирования и других подобных операций исключает возможность столь строгой соматотопичности. Однако неизбежная диффузность проекций еще не означает их неупорядоченности (само вычислительное устройство и не соматотопично, и не хаотично), даже если на сегодня мы не располагаем знаниями, которые бы дали возможность разглядеть закономерные структуры во взаимосвязях, скажем, лобной коры или миндалины.
* (Отметим четыре возможности уточнить эту схему: 1) включить в нее контуры обратной связи, в результате чего, например, механизмы устранения избыточности потенциальных команд смогут заставлять рецепторы добывать нужные данные; 2) попытаться показать, как некоторые функции, разделенные на этой схеме, могут переплетаться в различных структурах мозга; 3) включить в нее систему поощрений, первоначально управляемую генетически "запаянными" механизмами, которая может влиять на поведение и на выбор; 4) внести дополнительные контуры, показывающие, как ретикулярная формация может (согласно гипотезе, изложенной в разд. 7.1) воздействовать на общие направления выбора вида действий, закладывая тем самым фундамент для функционирования более тонких механизмов устранения избыточности потенциальных команд, намеченных в настоящей схеме.)
Мир состоит из разного рода объектов, и именно с объектами (или с их поведенческими признаками, или с группами объектов) нам приходится иметь дело. Поэтому, если мы хотим выжить в мире, в котором мы движемся относительно объектов, а объекты движутся относительно нас (см. разд. 5.5), наш мозг должен быть устроен таким образом, чтобы признаки, активированные стимулами от данного объекта, не только сохраняли активность до тех пор, пока этот объект остается важной частью среды, но и изменялись в соответствии с движением объекта относительно либо тех эффекторов, с помощью которых мы собираемся с ним взаимодействовать, либо тех рецепторов, с помощью которых мы регистрируем результаты этого взаимодействия.
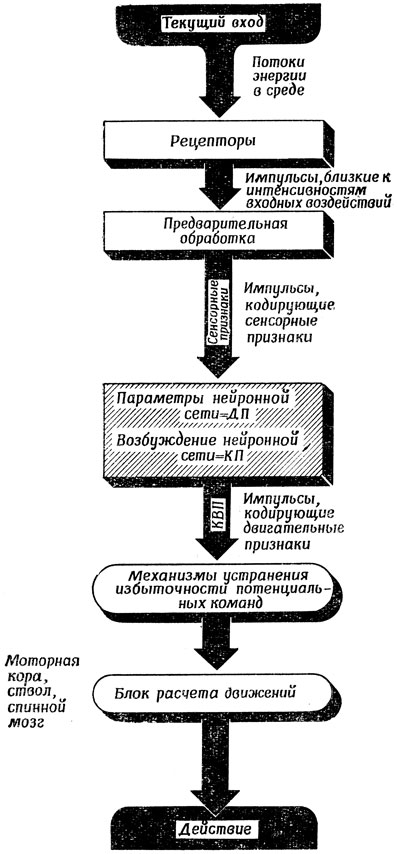
Рис. 103. Грубая блок-схема, напоминающая нам о функциях, участвующих в установлении взаимосвязи между прошлыми и настоящими входами, с одной стороны, и настоящим выходом - с другой (ср. с фиг. 109 и 117)
Последнее замечание станет более понятным, если мы сравним один из способов представления объектов в вычислительной машине с тем, как на основании опытов по исследованию реакции человека на вид вращающихся предметов можно представить себе характер аналогичного кодирования в биологических системах.
Прежде всего рассмотрим вкратце программу для вычислительной машины, предназначенную для ответов на вопросы, и противопоставим ей другой, по-видимому, более естественный с биологической точки зрения, гипотетический подход. Итак, как. составить программу для вычислительной машины, которая могла бы отвечать на вопросы типа "Мужчина ли президент Смит?", если в ее память введены различные кодированные высказывания о политической жизни США, и в частности о президенте Смите. Одно из таких высказываний может прямо утверждать: "Президент Смит-мужчина", и тогда машине остается лишь отыскать это утверждение, перебрав содержание своей памяти, после чего она сможет выпечатать: "Да, президент Смит - мужчина". Если же в памяти машины не содержится такого высказывания, то для получения ответа ей придется скомбинировать ответ из нескольких фактов, хранящихся в памяти. Например, там может найтись высказывание "Смит - президент" и "Все президенты были мужчинами". Найдя эти утверждения, машина сможет чисто формальным образом построить из них необходимый вывод: "И следовательно, президент Смит - мужчина", напечатав после этого ответ "Да".
В противовес такому чисто формальному подходу, который на сегодняшний день преобладает в работах по искусственному интеллекту, можно было бы сконструировать машину, которая использует вопрос "Мужчина ли президент Смит?" не для того, чтобы отыскивать фразы, относящиеся к политике и к президенту Смиту, и, комбинируя их, получать ответ, а использует имя "Президент Смит" для того, чтобы извлечь из памяти его фотографию, а затем найти программу распознавания образов, способную отличать фотографии мужчин от фотографий женщин, и поставить перед ней вопрос "Мужчина ли это?" Первый из намеченных подходов является синтаксическим, а второй - семантическим. И, по-видимому, человек, решая подобные задачи, использует оба эти подхода, а также много других.
Вводить в память картинку в виде двумерной матрицы слов, где каждое слово определяет интенсивность освещения соответствующей точки на картинке, а затем обрабатывать ее на нынешних вычислительных машинах, умеющих одновременно оперировать лишь с небольшим числом слов, - такой подход к задаче получения ответов на вопросы с помощью ЭВМ может оказаться весьма неэффективным. Однако, обратившись к мозгу, мы найдем в нем систему нейронов с практически трехмерным распределением функций, реагирующую на зрительное возбуждение. И так как в мозгу все клетки все время активны (в отличие от современных машин, в которых большая часть памяти совершенно пассивна), представляется вероятным, что мозг человека действительно может отвечать на такие вопросы, восстанавливая некоторое подобие пространственной нейронной структуры с высокой степенью распараллеливания на достаточно высоких уровнях нервной системы между входными путями и нейронами, несущими команды к мышцам, а затем уж обрабатывает эту возбужденную структуру точно так же, как если Оы она была вызвана зрительным стимулом. Впрочем, я хотел бы подчеркнуть, что такая возбужденная структура содержит входную информацию в закодированном виде, а никак не в виде простой "реставрации" зрительного входа. К этому замечанию мы еще вернемся в разд. 6.4, обсуждая голографическую метафору.
Серия опытов Шепарда и Метцлера дает нам некоторые сведения о том, каким образом человек различает разные виды внутренних представлений. Шепард предъявлял испытуемым объемные изображения объектов, которые можно сложить из кубиков (рис. 104). Испытуемому каждый раз показывали два объекта, которые были либо одинаковыми, либо представляли собой зеркальные отображения один другого. Если обе картинки изображали один и тот же объект, то одну из них получали из другой, как бы вращая объект либо в плоскости рисунка, либо вокруг оси, перпендикулярной этой плоскости. От испытуемого требовалось нажать одну из двух кнопок в зависимости от того, считал ли он, что на рисунках изображен один и тот же предмет, или нет.
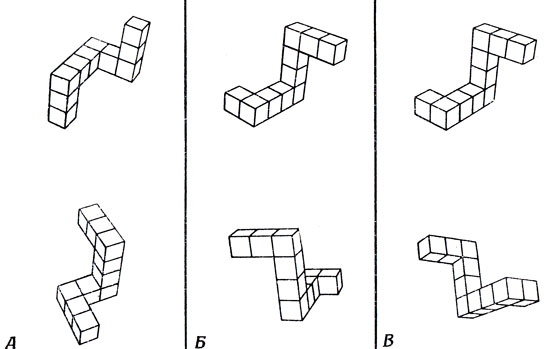
Рис. 104. Три примера (А-В) стимулов, использованных Шепардом в своих опытах. Каждый объект изображен в двух видах, причем второе изображение получено в результате вращения либо самого объекта, либо его зеркального отображения
Если подойти к этой задаче с позиций обработки вербальных описаний образов, то нам придется сначала генерировать описания типа "Здесь кубик, от него под таким-то углом отходит еще один, затем два прямых угла..." и затем снабдить каждое такое описание данными, указывающими на ориентацию описываемого объекта в пространстве. Если показать такой системе две картинки, то она проанализирует их, получит в результате такое описание, отбросит указания об ориентации и сразу приступит к проверке того, одинаковы эти описания или нет.
Другой подход состоит в том, чтобы, воспользовавшись двумерными картинками, создать в мозгу такую же картину возбуждения нейронов, какая получилась бы при разглядывании трехмерного объекта. В процессе реальных взаимодействий с трехмерными объектами мы привыкли поворачивать их, получая при этом их различные двумерные проекции. И можно предположить, что, поглядев на одну из таких двумерных проекций и восстановив "в уме" нейронное представление трехмерного объекта, мы окажемся в состоянии обеспечить нейронный эквивалент вращения объекта. (Не следует представлять весь этот процесс, хотя избежать этого трудно, слишком упрощенно и считать, что в мозгу строится объект из маленьких кубиков, который затем вращается, ведь мы говорим сейчас не о строго геометрических, а о функциональных соотношениях. Мы лишь хотим сказать, что в результате внутренних процессов в мозгу происходит преобразование картины возбуждения нейронов, родственное тому, которое было бы вызвано поворачиванием находящегося перед нами реального объекта. Мы уже допускали возможность таких операций, когда говорили о следящей составляющей в схеме на рис. 98.) Таким образом, мы можем представить себе, что испытуемый Шепарда вращает образ одного из показанных ему объектов и, накладывая полученный "мысленный образ" на образ другой картинки, решает, согласуются ли эти образы между собой или нет.
Испытуемые утверждают, что они действуют именно таким образом. При первых попытках они стараются проделать это слишком быстро и говорят, что к тому времени, когда они мысленно повернули объект, он "разваливается", они забывают, как он был устроен "на дальнем конце", и его приходится поворачивать снова. Но стоит им почувствовать, что они обучились делать это "с нужной скоростью", т. е. не забывают структуру объекта во время мысленного его вращения, как они начинают решать поставленную задачу безошибочно. Однако такой интроспективный подход уже не раз приводил к ошибочным результатам. Где же гарантия, что на этот раз он окажется надежным? В ответ на такое возражение Шепард решил измерять время реакции. Он рассуждал следующим образом. Если используется вербальный подход, то угловое смещение одного изображения относительно другого не должно сказываться на времени, нужном испытуемому для ответа. Если же испытуемый мысленно вращает картинку, то каждые новые 10° поворота должны добавлять какую-то определенную величину к суммарной задержке между моментом показа двух проекций и ответом испытуемого. Выяснилось, что между необходимым углом поворота изображения и числом секунд, затраченных испытуемым на обдумывание ответа, действительно имеется строго линейная зависимость, а это, по-видимому, подтверждает реальность "мысленного вращения изображения". Шепард установил, например, что он сам может поворачивать мысленный образ со скоростью 62,6° в секунду.
Описанный опыт служит примером интересных попыток разобраться в том, каким представляется нам внешний мир в нашем мозгу. Эволюция человека не готовила его к существованию в статичном двумерном мире. Мы перемещаемся в нашем мире, мы передвигаем и поворачиваем различные объекты, мы подходим к ним с разных сторон. И для того чтобы функционировать надлежащим образом, нам нужно уметь не только классифицировать объекты, но и соотносить их с собой, определять, где они расположены, понимать, как нам нужно передвинуть их или переместиться самим, чтобы надлежащим образом привязать эти объекты к эталонной системе координат, детерминированной нашим собственным телом. Именно поэтому столь важна мысль о том, что в мозгу имеются возможности не только для классификации или вербального описания, но и для воображаемого перемещения объектов относительно нашего тела или перемещения нас самих относительно этих объектов. Если же добавить к этому, что мозг тесно связан со всеми мышцами, в определенном смысле накладывающими на нас естественную систему координат, то мы увидим, что для определения поворота недостаточно, чтобы где-то в мозгу было записано соответствующее число. Это число должно быть, так сказать, всегда наготове, чтобы в любой момент привести в действие необходимые мышцы. Гвоздь опыта Шепарда заключается в том, что, измеряя время реакции испытуемых, он получил информацию, которую нельзя было получить лишь на основании того, верно или неверно они отвечали на вопросы. Обнаружив изящную линейную зависимость между необходимым углом вращения изображения и временем, затрачиваемым на выяснение разницы между объектами, Шепард позволил нам увидеть, на каких принципах основано хранение информации в мозгу. И одна из главных целей нашей теории мозга состоит в том, чтобы заставить морфологов и физиологов серьезно заняться поиском нейронных механизмов, обеспечивающих возможность подобного хранения.
Займемся еще одной группой данных, касающихся стимуляции мозга больных эпилепсией и указывающих на то, что сама организация мозга обеспечивает содержательное взаимодействие с внешним миром.
Приступая к операциям на мозге у больного эпилепсией, хирург должен представлять себе расположение различных областей мозга, с тем чтобы при удалении очага раздражения, вызывающего приступы эпилепсии, не повредить какую-либо жизненно важную структуру. Он раздражает электрическим током небольшие участки мозга и наблюдает реакции больного с целью выяснить, связан ли данный участок с управлением движениями руки, или маленького пальца ноги, или речи, или с чем-либо еще. Ведь если, например, в результате операции больной начнет слегка прихрамывать на левую ногу, то он, возможно, с радостью пойдет на такой компромисс; если же он рискует потерять речь, то вряд ли он согласится на операцию. Некоторые больные рассказывают, что при раздражении мозга они вновь "переживают" те или иные яркие эпизоды своей жизни. К сожалению, некоторые ученые считают это доказательством того, что в нашем мозгу хранится память обо всем, происходившем с нами в прошлом, и что к любому такому событию можно найти доступ, если достаточно постараться. Однако это предположение нелепо по многим причинам. Во-первых, в большинстве случаев мы не можем быть уверены, что больной говорит о действительных воспоминаниях. Ведь если его мозг создает какую-то конструкцию из "ингредиентов" прошлого опыта, то эта конструкция покажется ему похожей на что-то, что с ним уже случалось, хотя именно данной комбинации в прошлом могло и не быть. Во-вторых, нет никаких оснований рассматривать результат этих опытов как свидетельство в пользу "фотографической гипотезы", т. е. считать, что больной видит точно такую сцену, какую он видел в первый раз; вполне возможно, что конструкция, образовавшаяся в мозгу в результате раздражения, относится к достаточно высокому уровню нейронной активности, весьма далекой от реального зрительного входа. Наконец, подобные явления отмечались только у больных эпилепсией. Следует, однако, отметить, что, по их утверждению, такое "переживание" коренным образом отличается от "обычного воспоминания". Поэтому описанные факты свидетельствуют скорее о таком устройстве мозга, которое, в случае, если нам удастся проникнуть в достаточно высоколежащие центры и вызвать там искусственное раздражение, позволяет получить не группу необработанных фактов, поступивших на зрительный вход (скажем, не случайное сочетание света и тени), а нечто вроде осознанного восприятия объектов. Вводя электрод в мозг наугад и раздражая мозг электрическим током, мы вызываем в одной из нейронных сетей временную активность. И нет ничего удивительного в том, что такая временная активность часто приводит к тому, для чего эта сеть и предназначена, а именно к осознанному восприятию, а не к появлению на зрительном поле случайного рисунка световых пятен. (Быть может, это происходит не только у эпилептиков?) Нам кажется, что именно благодаря способности мозга освобождается от избыточности потенциальных команд случайный набор выходных признаков преобразуется в комплекс активности, связанной с ярким и более или менее реальным субъективным переживанием.
Итак, одним из центральных понятий нашей модели становится комплекс выходных признаков (КВП), или кодовое описание возможностей взаимодействия с объектом, содержащее набор признаков, необходимых организму для осуществления этого взаимодействия. (КВП представляет собой попытку воплотить понятие "диапозитив" из разд. 4.1 в несколько менее метафорической форме.) В то время как у низших животных взаимосвязь между сенсорными и двигательными признаками может быть в основном прямой и врожденной (ср. с функциями области тектума лягушки, разд. 7.2), у многих других форм существует дополнительный механизм, позволяющий совершенствовать эту взаимосвязь с учетом прошлого опыта животного. Таким образом, пока мы не делаем различия между генетически детерминированными механизмами и теми, которые образуются в процессе обучения, поскольку в процессе восприятия участвуют как те, так и другие, совместно создавая у животного долгосрочную модель окружающего мира.
Тем самым мы хотим подчеркнуть, что мы рассматриваем восприятие как представление окружающей обстановки в терминах открывающихся перед нами двигательных возможностей, а не в виде "маленькой копии мира", подобной фотографии.
Итак, мы выдвигаем гипотезу о том, что ДП (долговременная память) находится в нейронной сети, расположенной между сенсорными и двигательными слоями. Она сосредоточена в значениях параметров этой сети, позволяющей осуществлять разложение комплекса сенсорных признаков на элементы и преобразовывать их в набор КВП с данными о местоположении каждого объекта, что позволяет организовать взаимодействие с объектами, описываемыми теми сенсорными признаками, из которых были получены эти КВП.
Повторим еще раз, что такая система КВП, активированная в любой момент времени, очень часто содержит гораздо больше подпрограмм деятельности, чем организм может реализовать в этом момент; поэтому значительная часть нейронных механизмов должна заниматься тем, чтобы в каждый момент времени сосредоточить усилия организма лишь на одном наборе совместимых действий.
Кроме того, мы предполагаем, что КП (кратковременная память) системы охватывает всю текущую активность КВП. ДП обеспечивает организму возможность генерировать КВП по неполной (а возможно, и неоднозначной) сенсорной информации об объектах, тогда как КП обеспечивает сохранение соответствующих КВП в активном состоянии (хотя они и могут содержать лишь достаточно неопределенный контекст, на фоне которого происходит восприятие новых объектов), а не поставляет другие КВП, соответствующие лишь воспринимаемому в данный момент объекту. В разд. 4.4 мы уже высказали предположение (косвенно подтверждающееся данными Шепарда), что мозг располагает возможностью непрерывно переадресовывать или видоизменять КВП, с тем чтобы их расположение все время соответствовало представляемым ими объектам, даже если такая переадресовка относительна и связана с изменением направления входных (сенсорных) и выходных (двигательных) путей. Так, если мы кружимся вокруг стола, движения руки, необходимые для того, чтобы взять лежащий на столе карандаш, непрерывно меняются, а если мы отойдем от стола, то к выходным признакам карандаша добавятся еще и такие "фоновые признаки", как необходимость вернуться к столу для того, чтобы получить возможность добраться до карандаша. "Конечно" (вот слово, вызывающее обычно тысячи вопросов!), в сознание взрослого человека такие ориентированные на действие признаки попадают лишь в вербальной форме (например: "Это находится в двух метрах от меня"). Поэтому нервная сеть должна быть устроена таким образом, чтобы в общем случае было легче обнаружить согласованность возбужденного КВП и комплекса сенсорных признаков, несмотря на несоответствие в расположении ("А вы переместились"), и переадресовать КВП нужным образом, чем с помощью ДП заново генерировать весь КВП ("Кто вы?"), не имеющий прямого отношения к текущим входным признакам. Иными словами, КП и внимание "переплетены" друг с другом: мы обращаем внимание лишь на то, что меняется (или на то, для чего мы считаем изменение неизбежным и важным), и это отражается на всех слоях нервной системы и на всех процессах обработки сенсорной информации.
Подчеркнем поэтому еще раз, что КВП - это картина нейронного возбуждения, отвечающая некоему объекту с точки зрения одного или нескольких возможных взаимодействий организма с этим объектом. (Один и тот же объект может вызывать различные КВП в разных ситуациях. Так, кошка может возбудить выходной признак "гладить" в одном случае и "прогнать" - в другом. Можно сказать, что преобразование от объекта через посредство сенсорных признаков к КВП приводит к политетической классификации.) Статус КП как совокупности данных о том, что можно делать с окружающей средой, можно считать эквивалентным внутренней модели среды, близкой нам с точки зрения пространства, времени и значимости для нас. Очевидно, что она гораздо полезнее для организма, чем простое копирование текущей сенсорной стимуляции.
Таким образом, КВП - это не просто копия в (случае зрительного восприятия) некоторой двумерной картинки. Например, на рис. 105 показаны два изображения куба, создающие различные наборы сенсорных признаков, но которые могут тем не менее вызывать одни и те же КВП, связанные, например (может быть, наряду со многим другим), с возможностью взять этот куб в руку, с точностью до различий в параметрах, отражающих разницу в величине угла между большим пальцем и ладонью при попытке взять в руку настоящий куб, который бы соответствовал одной из этих двумерных проекций.
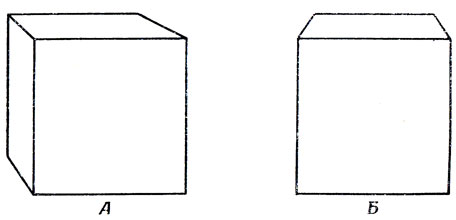
Рис. 105. Два изображения, которые нельзя перевести друг в друга движениями в плоскости. У одного 7 вершин и 9 ребер (А), а у другого 6 вершин и 7 ребер (Б). Однако это расхождение по сенсорным признакам отбрасывается, как только мы распознаем, что и то и другое - куб
Опираясь на все эти рассуждения, мы можем теперь перейти к описанию схемы, позволяющей преобразовывать как сенсорную входную информацию, так и моторный выход уже после КВП, с тем чтобы обеспечить плавное взаимодействие организма с окружающим его миром.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'