6.3. Преобразования и соматотопическая организация
Теперь наша основная задача состоит в том, чтобы в схему на рис. 103 "ввести" соматотопию, привязав ее к нашим соображениям о распределенной системе управления движением (разд. 5.5) и показав, где могут производиться некоторые преобразования комплексов сенсорных признаков и наборов КВП, прошедших предварительную переработку, с тем чтобы обеспечить надежное взаимодействие организма с окружающим миром.
Прежде всего следует подчеркнуть, что имеется достаточно оснований допустить наличие у млекопитающих двух (в высшей степени независимых) зрительных систем, из которых одна связана с корой, а другая - со средним мозгом. Различие между этими системами становится понятным, если вспомнить опыты Шнейдера [227] на хомяках: когда у хомяка разрушают зрительную кору, не затрагивая верхнее двухолмие, то он теряет способность различать даже простейшие зрительные образы, например не отличает квадрат от креста, хотя по-прежнему может ориентироваться на звук или на движущийся предмет. Другими словами (ср. противопоставление кошки и лягушки в разд. 2.4), создается впечатление, что хомяк без зрительной коры все еще может реагировать на простейшие локальные признаки, особенно если они связаны с движением, но не способен интегрировать их в глобальные детерминанты КВП. И наоборот, если у хомяка удалить верхнее двухолмие, не затрагивая зрительной коры, то он различает образы, но не в состоянии использовать обнаруженные различия для ориентации. На основании этого последнего наблюдения многие исследователи делали вывод, что хомяки с ненарушенной зрительной корой и удаленным верхним двухолмием просто слепы: когда такого хомяка помещают в установку, изображенную на рис. 106, то он вроде бы с равной вероятностью направляется как в тот отсек, где за дверью (помеченной квадратом) его не ждет никакое вознаграждение, так и в тот отсек, где за дверью (помеченной крестом) он получит вознаграждение (воду). Однако Шнейдер обратил внимание на то, что, хотя животное и может войти в "не тот" отсек, оно научается не входить в "не те" двери и, дойдя до них, поворачивает назад. (Шнейдер исключил возможность близорукости животных, поставив стеклянные двери и поместив опознавательные знаки на некотором расстоянии за ними. Это не мешало животному научиться распознавать их.) Полученные данные свидетельствуют о том, что животное с интактной корой, но без верхнего двухолмия способно различать образы, но не в состоянии "привязать" их к месту. Оказалось, что такое животное не умеет ориентироваться и по "локальным признакам": если ему предъявляют звук или движущийся предмет, оно "застывает" на месте, а не поворачивается к источнику стимуляции, как это сделало бы животное, не лишенное верхнего двухолмия.
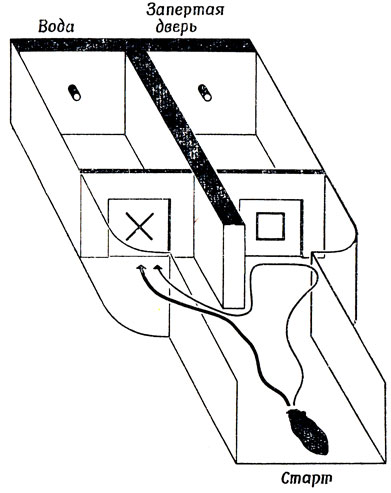
Рис. 106. Интактный хомяк (траектория его движения показана жирной линией) может ориентироваться по зрительным стимулам. После удаления верхнего двухолмия хомяк, по-видимому, не может использовать зрительные признаки для ориентации, но по-прежнему способен различать их; об этом свидетельствует траектория его движения (тонкая линия), показывая, что животное руководствуется тактильным ощущением, но толкает только ту дверь, на которой имеется знак, ассоциирующийся у него с вознаграждением
С учетом всех этих соображений перейдем к рис. 107, где приведена грубая блок-схема системы, содержащей многие аспекты "стратегий млекопитающих", и в частности все только что высказанные соображения и многое другое, о чем мы еще поговорим ниже, рассматривая эту схему во всех ее подробностях. Конечно, эти же самые функции могут быть реализованы и многими другими структурами. Кроме того, мы увидим, что, несмотря на многочисленность разных блоков в нашей системе, она тем не менее отражает организацию деенаправленного восприятия лишь в самом первом приближении. Мы надеемся, однако, что, изучая эту схему, мы сможем вплотную подойти к следующему, более совершенному уровню рассмотрения восприятия и, возможно, выдвинем новые гипотезы, которые помогут нам связать отдельные функции восприятия с соответствующими структурами мозга.
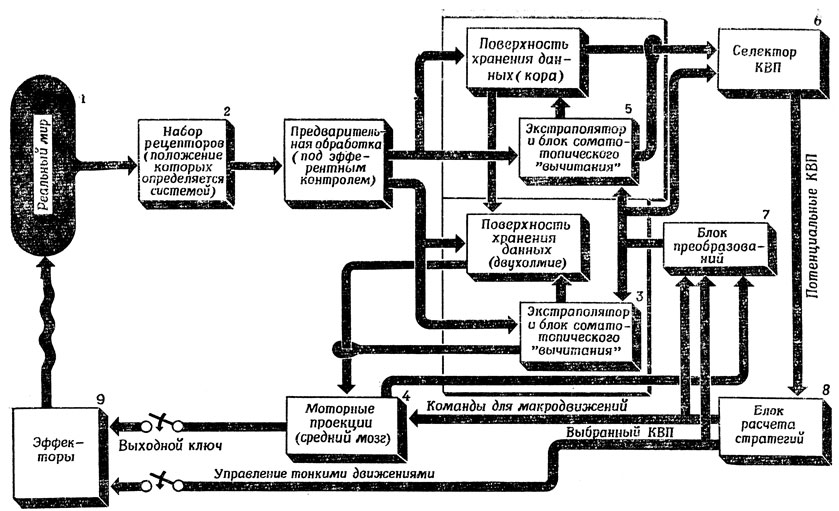
Рис. 107. Предположительная блок-схема взаимодействия 'двух зрительных систем' среднего мозга (3) и коры головного мозга (5), обеспечивающая сохранение правильных взаимосвязей между организмом и внешним миром
Можно считать, что объекты реального мира (1) оказывают влияние на потоки энергии в окружающей среде. Различные системы рецепторов животного (2) регистрируют проекции этих потоков на различные сенсорные модальности и различные участки пространства. Зрительные рецепторы, тактильные рецепторы рук и т. п. можно перемещать относительно остальных частей тела, и эти перемещения частично соответствуют влиянию, которое оказывает на эффекторы блок 4 (см. ниже). Входные сигналы, полученные с помощью рецепторов, проецируются в две основные системы: одну в среднем мозгу (3) и другую - в коре полушарий (5). Попадая в средний мозг, эти сигналы подвергаются локальной (поточечной) предварительной переработке и попадают в механизм того же типа, что и показанный на рис. 101. Области, получающие стимуляцию, которая не совпадает с ожидаемой, вызывают движения ориентации, для чего в работу включаются моторные структуры (4). Питтс и Мак-Каллок [202], описавшие эту схему, считают, что именно таким образом глаз приводит рассматриваемый объект в "стандартное положение" на зрительной коре, где осуществляется распознавание, и таким образом действует через эффекторы, обеспечивая правильное расположение в пространстве входного сигнала, поступающего в кору.
Переходя на кортикальные пути, сенсорные сигналы попадают в другой механизм типа показанного на рис. 101 (5); этот механизм пропускает в селектор КВП (блок распознавания образов, генератор гипотез об объекте, 6), занимающийся пространственным анализом картины возбуждения, лишь те участки входного образа, которые подвержены изменениям во времени, сильно отклоняющим их от прогноза, основанного на учете собственного движения организма или подмеченного движения объектов. Эта схема работает потому, что те же области непредвиденных изменений (с помощью механизмов ориентации 3 я 4) заставляют, как уже отмечалось выше, рецепторную поверхность поворачиваться поочередно к каждому из этих источников новой информации. Селектор КВП получает также сведения о том, к чему приводят текущие собственные движения, получая их из блока 7, в результате чего выбор КВП согласуется с текущей активностью. Решения, принимаемые с оглядкой на "цели организма" (увы, "флогистон" наших нынешних рассуждений), позволяют выбрать один из потенциальных КВП, генерируемых блоком 6. Блок вычисления стратегий (5) реализует затем выбранный КВП, а управление тонкими движениями, накладываемыми на макродвижения, осуществляется в рамках естественной системы координат, детерминируемой нашим телом и поддерживаемой средним мозгом.
Приняв такую схему, мы можем истолковать результаты опытов Шнейдера с хомяками как свидетельство того, что при разрушении зрительной коры (что соответствует отключению блока 5 нашей схемы) животное сохраняет способность ориентироваться в пространстве (способность локализации), но не способно к распознаванию образов, а при разрушении верхнего двухолмия (блок 3 на нашей схеме) животное способно распознавать образы, если его взгляд случайно упадет на объект именно так, как это необходимо для получения стандартного возбуждения КВП, но не способно к необходимым ориентирующим движениям.
Представьте себе, что человеку показывают проекцию диапозитива незнакомой ему сцены, причем диапозитив вставлен в проектор, скажем, "вверх ногами". В этом случае испытуемому придется провести весьма трудоемкий анализ картины. Прежде всего ему нужно понять, что все, что он видит, перевернуто "вверх ногами". Затем ему придется либо изменить положение своих рецепторов (2), например встав на голову, либо произвести сдвиг в выходной моторной проекции (4), это будет осуществлено преобразователем (7), который перестроит параметры селектора КВП (6). Затем придется осуществить и другие преобразования, направленные на то, чтобы привести изображение к стандартной форме. Все это вызовет такое изменение положения рецепторов, моторных проекций (и, возможно, положения эффекторов) и функций предварительной переработки, что система восприятия "подстроится" к той гипотезе о КВП, которая проверяется в настоящее время. А это значит, что блоки, которые мы столь легкомысленно окрестили "селектором КВП" (6) и "блоком вычисления стратегий" (8), должны быть весьма сложны и тесно связаны с другими частями системы.
Сосредоточим теперь наше внимание на том аспекте восприятия, который за недостатком места не включен в рис. 107. Для этого мы обратимся к рис. 108, на котором приведена схема, обобщающая схему Питтса - Мак-Каллока [202] (рис. 96).
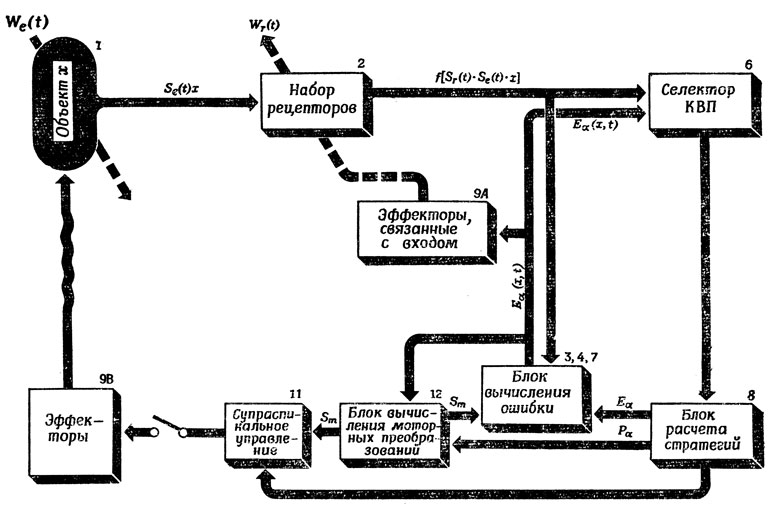
Рис. 108. Подробности того, как модифицированная схема Питтса - Мак-Каллока (рис, 96) может заменить регулятор для селектора КВП на рис. 107,
Для того чтобы облегчить переход от рис. 107 к рис. 108, заметим, что для простоты блок 5 на рис. 108 опущен и что блоки 3 и 4, а также преобразователь (7) считаются блоком вычисления ошибки, а их воздействие на селектор КВП показано линией, соединяющей на рис. 108 блок вычисления моторных преобразований (12) с блоком вычисления ошибки (5, 4, 7). На схеме 108 появился еще и блок супраспинального управления (11), соответствующий многоэтапным преобразованиям КВП в сигналы управления отдельными мышечными волокнами.
Другими словами, на схеме 108 мы осуществили декомпозицию механизмов формирования и выбора КВП на подсистемы, в явном виде участвующих в поиске соответствующих преобразований и в управлении движением с помощью параметризации КВП, отделив их от остальных блоков схемы. Одновременно мы замаскировали на этой схеме анатомическую обособленность двух различных путей, показанных на рис. 107. Рассматривая схему 108, нужно иметь в виду следующее. Схема 107 в неявном виде предполагает, что система всегда как бы воспринимает изображение в том смысле, что КВП, вызывающий решение типа "Обойдем-ка вокруг этого предмета, и, возможно, нам удастся понять, что же это такое", можно расценивать как восприятие объекта, как чего-то такого, вокруг чего можно ходить. На рис. 108 эти первые поисковые фазы настройки движения и восприятия выделены и в явном виде объясняется, как эти процессы приводят к более "полному" восприятию объекта ("Ах, это же вверх ногами, это фотография памятника Вашингтону"), позволяющего назвать его.
Реакцию ориентации не следует при этом рассматривать как один из входных признаков объекта, хотя она и оказывает влияние на пространственное расположение КВП относительно соматотопической системы координат КП. Ориентация делает возможным формирование тонких признаков КВП и, следовательно, предшествует такому формированию или по крайней мере его уточнению. Для животных с фовеальным зрением, т. е. для тех, в глазу которых имеется центральная ямка (fovea), происходящее в коре "распознавание образов" можно считать организованным по принципу "распределения времени" (если вновь прибегнуть к метафоре, заимствованной из вычислительной техники); механизмы ориентации и необходимая экстраполяция, осуществляемая в коре, определяют местоположение блоков "распознавания образов", как если бы выяснение того, что же здесь имеется, требовало более сложной "обработки", чем обнаружение несоответствия с ожиданием. Таким образом, человек оказывается где-то посредине между лягушкой, у которой совсем нет центральной ямки и обработка зрительного входа осуществляется равномерно и в относительно ограниченных масштабах, и неким "сверхчеловеком", у которого весь глаз - сплошная центральная ямка и который обрабатывает равномерно и весьма совершенным образом огромное зрительное поле.
Положение и ориентация объекта относительно рецепторных поверхностей организма частично определяют, какие потоки энергии из внешней среды достигнут этих поверхностей. Назовем некоторое положение объекта стандартным и обозначим его через л;. В любой момент времени t его положение в абсолютном пространстве может рассматриваться как результат его переноса из x и соответствующего вращения. Если обозначить суммарный эффект преобразований через Se(t), то возникшее распределение вещества в пространстве можно будет описать выражением Se(t)•x. Положение рецепторных поверхностей в момент времени t обеспечит проецирование этого распределения, которое можно описать оператором Sr(t), так что нейронное представление объекта можно будет рассматривать как некоторую функцию
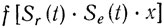
двумерного образа. (Мы говорим сейчас исключительно о зрительных рецепторах, однако с небольшими изменениями все эта приложимо и к другим модальностям.)
Конечно, взрослый человек способен мыслить в абстрактных категориях или планировать свои движения "извне", как бы представляя, что он передвигается по карте. Но нам хотелось бы здесь ограничиться рассмотрением основной "эгоцентрической" формы восприятия, при которой организм представляет объекты не в терминах какой-то абсолютной системы пространственных координат, а в зависимости от их положения относительно того, кто их воспринимает, т. е. в терминах системы координат, определяемой тем, насколько далеко может достать его рука, в какую сторону он повернулся лицом, числом шагов до объекта и т. п. Б этом случае размеры объекта будут восприниматься в зависимости от масштаба движений, необходимых для желаемого взаимодействия с объектом, ориентация - от того, насколько нужно повернуть голову для того, чтобы сфокусировать взгляд на объекте, и т. п. Например, можно утверждать, что мы воспринимаем линейки на листе линованной бумаги не только в той мере, в какой мы можем попасть на них кончиком карандаша, но и в той, в какой нам удастся приспособить свой почерк к расстоянию между линейками. Иначе говоря, наша теория должна учитывать еще и преобразование Sm(t), приводящее мышечную деятельность к масштабу, - соответствующему решаемой задаче.
Учитывая важнейшую роль, которую играет в нашей теории порождение преобразований, обеспечивающих согласование моторного выхода и сенсорного входа, мы в состоянии теперь оценить, насколько важно для нас описанное выше обобщение (см. рис. 96) "принципа однородности конструирования рефлекторных механизмов, обеспечивающих инвариантность относительно произвольной группы G преобразований входных воздействий", впервые предложенного Питтсом и Мак-Каллоком [202]. Однако если в исходной модели рассматривается лишь возможность преобразования сенсорных признаков (например, возможность вращения зрительной перспективы в плоскости), направленного, скажем, на то, чтобы центрировать сенсорный стимул на зрительном поле, то нас интересуют и преобразования выходных признаков, соответствующие всевозможным перемещениям объекта в пространстве; ведь организму необходимы данные (хотя, как выяснится впоследствии, не все они обязательно должны быть закодированы в нейронной сети) об относительном расположении объектов, если только он собирается взаимодействовать с ними. (Питтс и Мак-Каллок рассматривали случай, когда такое преобразование реализуется движением глаз, но мы допускаем и возможность внутренней "переадресации" возбуждения.) Мы хотели бы, чтобы двигательные возможности организма позволяли осуществлять преобразования, приводящие к такому изменению параметров КВП, в результате которого наши пальцы будут смыкаться вокруг объекта, а не хватать воздух рядом с ним.
Так или иначе, напомним, что в исходной схеме с каждым образом φ связывался некоторый n-мерный "вектор ошибки" E(φ), обладавший тем свойством (ниже мы сможем существенно ослабить это требование), что E(φ)=0 тогда и только тогда, когда φ приведено к стандартной форме, и некоторое, зависящее от выбранной меры ошибки E(φ) преобразование W позволяющее эту ошибку уменьшить. Другими словами, мы выбирали для этой схемы такое W:Rn→G, что для всех образов φ
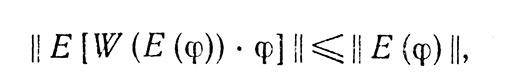
причем это неравенство заменяется на равенство тогда и только тогда, когда φ приведено к стандартной форме.
Мы уже имели случай отметить сходство этой схемы с методом операторов из ОРЗ (общего решателя задач) Ньюэлла, Шоу и Саймона [184] и два важных различия между ними: 1) в ОРЗ предполагается, что число разных "векторов ошибки" или "различий" конечно, 2) оператор WD, соответствующий различию D, не обязательно должен уменьшать это различие. В связи с последним свойством ОРЗ возникает потребность в особой процедуре, известной как эвристический поиск (разд. 4.2); смысл ее состоит в том, чтобы найти последовательность операторов (среди множества по внешнему виду подходящих последовательностей), которая действительно устранит исходное различие. Именно эту, далеко не идеальную, но гораздо более реалистическую ситуацию мы будем иметь в виду, когда прибегнем ниже к эвристическому поиску. Изменение We(t) расположения объекта может быть вызвано как его автономным движением (1), так и воздействиями на него моторного аппарата (9 В). Изменение Wr(t) расположения набора рецепторов осуществляется связанными с входом эффекторами (9 А), которые действуют как и в схеме Питтса - Мак-Каллока, на основе вектора ошибок, вычисляемого в соответствующем блоке (3, 4, 7). Аналогично блок вычисления моторных преобразований (12) вырабатывает задания для блока супраспинального управления (11), основываясь на тех же данных блока вычисления ошибки.
Однако, обсуждая работу блока (6), мы наталкиваемся на дополнительные тонкости, в связи с чем возникает своего рода дилемма "курицы и яйца". Трудность здесь состоит в том, что мы не можем решить, какая форма входного образа является стандартной, пока не знаем, что же представляет собой воспринимаемый объект, а в то же время привести воспринимаемый образ к стандартному виду необходимо именно для идентификации этого объекта. Выше мы выдвинули гипотезу о том, что распознавание объекта системой равносильно появлению у нее подходящих (с точки зрения целей организма, определяемых блоком расчета стратегий) КВП, обеспечивающих возможность взаимодействия с объектом. С каждым из таких КВП мы связываем какой-то критерий ошибки: каждый КВП считается пригодным для взаимодействия с объектом, лежащим за данным входным возбуждением, только тогда, когда этот входной образ приведен к такому виду, при котором некоторая фиксированная компонента E1 ( в этом и состоит ослабление требований для схемы Питтса - Мак-Каллока, которое мы обещали выше) вектора ошибки достаточно близка к нулю, в этом случае остальные компоненты E2 вектора ошибки служат параметрами для выполнения соответствующей программы. Таким образом, критерий ошибки должен определяться как текущими целями системы, так и имеющимися у нее оценками того класса объектов, к которому принадлежит правильно распознанный объект.
Селектор КВП (6) выполняет нечто подобное эвристическому поиску, хотя, впрочем, речь здесь идет скорее о поиске критерия ошибки, чем об уменьшении ошибки. Выдвинув гипотезу о том, что адекватный КВП для управления взаимодействием с объектом - это Pα, этот блок должен сообщить блоку вычисления ошибки (5, 4, 7) критерий точности Eα, а последний блок должен сообщить блокам (9А) и (12) значения вектора Eα(x,t), используемые в них для коррекции расположения рецепторов Sr(t) и моторного преобразования Sm(t). При этом блок (6) следит за вызванной этим изменением ошибкой и решает, насколько удачен или неудачен текущий выбор Pα. Предположим, что выбор Pα по отношению к имеющейся внешней среде достаточно оправдан и что в результате контрольная компонента E1α(x,t) становится достаточно близкой к нулю, что свидетельствует о возможности применения предлагаемой программы Pα. Понятно, что, как и в процедурах эвристического поиска, нам придется предположить существование некоторого правила, позволяющего судить, в какой момент невозможность свести E1α к нулю должна указать на необходимость поиска новой гипотезы.
Блок расчета стратегий разрешает выполнение некоторого двигательного действия только в том случае, если E1α(x, t) достаточно мало. В этом случае он запускает "интерпретирующую" подпрограмму, определяемую текущим выбором программы Pα и значениями параметров, задаваемыми остальными компонентами E2α(x,t) вектора ошибки. Основываясь на этой подпрограмме и информации, заключенной в векторе ошибки, блок моторных преобразований (12) определяет необходимую информацию о масштабах Sm(t), причем ее адекватность выясняется с помощью контура обратной связи, замыкающегося в блоке вычисления ошибки, и проявляется в сведении E1α(x, t) к нулю. Наконец, блок супраспинального управления (11) интерпретирует "подпрограмму", выбранную для него блоком расчета стратегий (5), используя при этом информацию Sm(t), поступающую в него из блока моторных преобразований (12).
Отметим, что описанная схема не требует хранения в нервной системе всей информации о преобразовании T, как это необходимо для того варианта схемы Питтса - Мак-Каллока, который показан на рис. 96, и допускает движения моторного аппарата, вызывающие изменения относительной проекции входного образа, в связи с чем любые абсолютные преобразования теряют смысл. Например, поворачивая голову так, чтобы все время смотреть на мышь, кошка одновременно настраивает свой моторный аппарат на прыжок. И главное здесь в том, что мозг не должен хранить данные о всех возможных абстрактных преобразованиях, а вместо этого выполняет операции, которые обеспечивают взаимосвязи между организмом и объектом, необходимые для их возможного взаимодействия.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'