7.2. Визиомоторная активность лягушки
Для того чтобы противопоставить что-нибудь модели ретикулярной формации и придать конкретность нашим соображениям о многослойном вычислительном устройстве (гл. 6), мы построим на основе принадлежащей Питтсу - Мак-Каллоку модели управления взглядом (разд. 5.5) соматотопически организованную модель управления тем поведением лягушки, в. основе которого лежит зрительная информация.
Сначала мы расскажем вкратце о некоторых аспектах визиомоторной деятельности лягушки, а затем ограничимся рассмотрением тех из них, которые укладываются в нашу модель.
Лягушка может сидеть неподвижно несколько часов подряд, но если в ее поле зрения появляется небольшой трепещущий объект, она может отреагировать на это либо ориентировочной реакцией (т. е. повернуться в сторону этого объекта), либо реакцией нападения (т. е. повернуться к нему, "выстрелить" языком и схватить его). Ингл установил, что для лягушки существуют две четко выраженные зоны, определяемые относительно положения ее головы, а не тела; если трепещущий объект попал во внутреннюю зону, то лягушка нападает на него, а если во внешнюю, то только поворачивается к нему.
В поведении лягушки особенно интересно отметить два момента, хотя мы и (не будем пытаться моделировать их. Во-первых, движения лягушки организованы по баллистическому принципу: она как бы "стреляет" по цели, не имея никакой возможности корректировать свои действия после того, как они начались, поскольку, если экспериментатор удалит из поля зрения лягушки стимул в течение приблизительно первых 100 миллисекунд после того, как она начнет движение, лягушка все равно "ударит" по исходной позиции стимула. Во-вторых, лягушка нападает на "муху", даже если эта муха попала в поле зрения только одного глаза, так что бинокулярное зрение для этого, по-видимому, не требуется. (Ингл предположил, что аккомодация глаза, необходимая для точной фокусировки стимула, дает лягушке необходимую информацию о расстоянии до цели, но обнаруженные Китингом и Гейзом [135] интертектальные связи, которые способствуют бинокулярному зрению, наталкивают на мысль, что при более тщательных исследованиях удастся обнаружить какой-то специальный механизм "тонкой настройки", включающийся в работу только тогда, когда оба глаза действуют согласованно.)
В настоящем разделе нас будет прежде всего интересовать поведение лягушки в присутствии более чем одной "мухи". В подобной ситуации вероятность активной реакции лягушки уменьшается, а если реакция и происходит, то лягушка обычно нападает лишь на одну из "мух". Однако Ингл [124] установил, что в ситуации, показанной на рис. 113, когда оба стимула движутся синхронно и симметрично, то приближаясь к носу лягушки, то удаляять от него, она обычно "бьет" в точку, находящуюся строго посередине между этими стимулами. Можно сказать, что она нападает на "среднюю" муху. Наша задача состоит в том, чтобы построить нейронную сеть, которая, используя информацию о "пищеносности" среды, соотнесенной с пространственными координатами и полученной либо с помощью "детекторов насекомых" (тип II, см. разд. 2.4), либо от нейронов тектума, комбинирующих информацию от этих детекторов с сигналами, полученными из других слоев, обеспечит воздействие на управление двигательной деятельностью системы лишь одного источника возбуждения.

Рис. 113. Лягушка, нападающая на 'среднюю муху'
Наши соображения, изложенные в разд. 6.1, заставляют нас искать для этой задачи решение, основанное на распределении вычислительных функций. Поэтому прежде всего мы вкратце перечислим те стратегии, от рассмотрения которых мы отказываемся. Одна из таких стратегий - это последовательное сканирование, суть которого иллюстрирует рис. 114: животное под действием какого-то центрального органа последовательно сканирует "предполагаемую зону пищеносности" до тех пор, пока не наткнется на область высокой активности; после этого оно прекращает сканирование, считывает текущие "координаты сканирования", отдает соответствующий приказ системе управления движением и в результате всего этого "бьет" в соответствующую точку пространства. Подобная схема может, вероятно, объяснить даже "эффект средней мухи", если предположить, что в каждом глазу имеется свое сканирующее устройство, так что в случае симметричных стимулов оба процесса сканирования прервутся строго одновременно, а противоположные движения в ответ на противоречивые команды приведут к "среднему" движению. Но несмотря на всю привлекательность подобного механизма сканирования, мы должны отказаться от этой гипотезы, ибо, не говоря уже о наших общих соображениях в пользу соматотопической организации управления, этот механизм потребовал бы слишком много времени на сканирование того множества объектов, которые попадают в поле зрения лягушки.
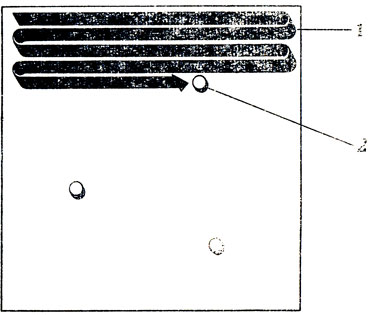
Рис. 114. Небиологичный способ устранения избыточности потенциальных команд, основанный на последовательном сканировании. 1 - путь сканирования; 2 - первая встреченная область высокой активности
В поисках схемы, которая использовала бы пространственное распределение информации, мы обратимся к работе Диддея [59]; в ней предложена новая структура, основанная на конкурентном взаимодействии разных частей тектальной области, в результате которого в обычных обстоятельствах лишь наиболее активная из этих частей передает на систему двигательного управления входной сигнал выше порогового; при этом предполагается, что система управления движением имеет некоторую пространственно распределенную входную поверхность, подобную той, которая изображена на рис. 99. Мы начнем с того, что введем новый слой клеток, соответствующий "слою пищеносности" и предназначенный для генерирования входов для системы управления движением. Таким образом, в определенном смысле характер возбуждения, приходящего в данную клетку из соответствующего рецептивного поля, должен описываться слоем "относительной", а не "абсолютной пищеносности".
В предложенной Диддеем схеме преобразования "пищеносности" в "относительную пищеносность" имеется популяция так называемых клеток сходства, каждая из которых как бы "контролирует" целый участок "слоя относительной пищеносности" и "приглушает" активность, которую клетки этой группы получают от соответствующих клеток "слоя абсолютной пищеносности", тем сильнее, чем выше активность клеток, лежащих за пределами контролируемого района (рис. 115). Этот механизм гарантирует, что высокая активность в каком-либо участке "слоя абсолютной пищеносности" передается "слою относительной пищеносности" только в том случае, если этот участок не окружен областью столь же высокой активности, которая могла бы блокировать эту передачу.
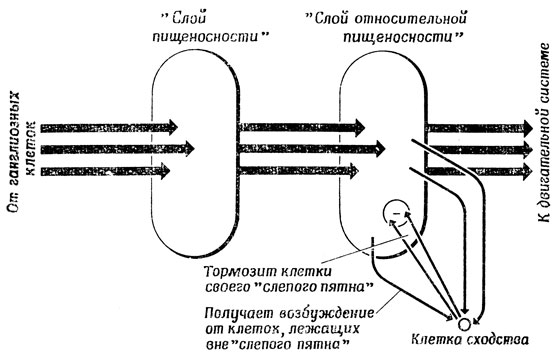
Рис. 115. Функционирование клеток сходства
Исследуя поведение подобной системы, мы выясним, что правдоподобная схема соединения указанных слоев должна обладать следующими свойствами:
1. Если активность в каком-либо одном участке выше активности в любом другом участке, то этот участок в конце концов "возьмет верх" над всеми остальными и животное ударит по соответствующей точке пространства.
2. Если два разных участка обладают достаточно близкой активностью, то
а) если оба участка обладают весьма высокой активностью, то они оба могут "взять верх" на другими участками входной поверхности и одновременно "взять на себя" управление, в результате чего лягушка будет бить по точке, лежащей посередине между этими двумя областями;
б) однако во многих случаях эти два активных участка просто "погасят" эффект друг друга, в результате чего ни один из них не сможет взять на себя управление и лягушка останется неподвижной, не обращая внимания ни на одну из "мух".
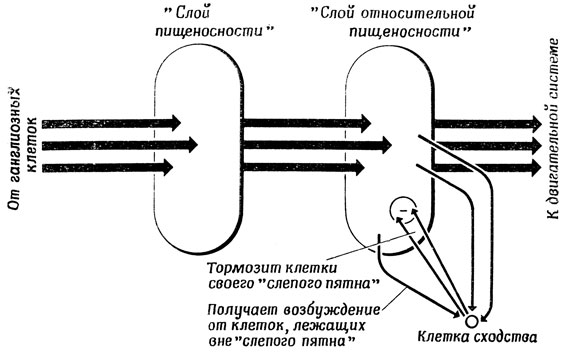
Рис. 116. Функционирование клеток новизны. Клетки новизны вносят временные изменения, происходящие в 'слое пищеносности', в вычисления 'слоя относительной пищеносности'
Одно из затруднений в этой схеме состоит в том, что нарастание торможения, создаваемого клетками сходства, исключает возможность быстрой реакции системы на новый стимул. Например, если бы в случае 2,б один из двух участков очень высокой активности внезапно стал еще более активным, то казалось бы, что образовавшийся затор должен очень быстро ликвидироваться. Однако для описанной выше системы это не так из-за наработанного торможения в клетках сходства, контролирующих этот участок. Поэтому Диддею [59] пришлось ввести на каждую клетку сходства по одной клетке новизны. Задача клеток новизны состоит в том, чтобы следить за изменениями активности соответствующего участка во времени. Если такая клетка обнаруживает достаточно резкое повышение активности в контролируемом участке, то она перекрывает тормозное воздействие клетки сходства и позволяет перенести новый уровень активности в слой относительной пищеносности (рис. 116). Эта модификация позволяет преодолеть инерционность старой модели, и таким образом мы получаем "мозг лягушки", который быстро реагирует на новые содержательные стимулы (рис. 117).
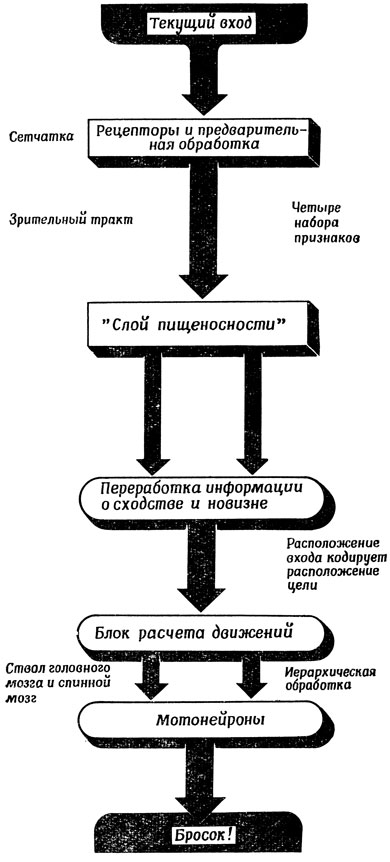
Рис. 117. Общая схема визиомоторной активности лягушки. Схема сходна с рис. 103 и 109 с той разницей, что в этой простой модели не предусмотрено использование долговременной памяти для модификации сенсорного входа
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'