7.1. Модель ретикулярной формации
В этом разделе мы рассмотрим модель, предложенную Килмером и Мак-Каллоком, которые пытались объяснить, каким образом РФ может управлять выбором типа поведения организма. Однако сначала мы познакомимся с некоторыми данными по анатомии нервной системы, на основе которых Килмер и Мак-Каллок строили свою модель. М. Шейбл и А. Шейбл, изучая срезы ствола головного мозга десятидневных крыс, подобные показанным на рис. 111, А, заметили, что коллатерали, идущие как от ретикулярных аксонов клеток РФ (верхняя часть схемы), так и от пирамидного пути (рис. 111, 3), располагаются в основном в плоскостях, примерно перпендикулярных главной оси ствола. Дендриты клеток РФ также, по-видимому, распределены по таким плоскостям, чем они резко отличаются, например, от дендритов ядер подъязычного нерва (рис. 111, 1). В связи с этим указанные авторы выдвинули гипотезу, согласно которой ретикулярную формацию можно рассматривать как стопку "фишек" - "сегментов" нейропиля (например, нейронного "войлока), как это показано на рис. 111, Б.
![Рис. 111. Схема строения ствола мозга. У типичного нейрона ретикулярной формации аксоны направлены как каудально, так и краниально, а дендриты ветвятся по поперечным плоскостям (А). Это навело на мысль, (см., например, [223]) о том, что РФ можно упрощенно представить себе в виде стопки игральных фишек (Б). 1 - ядро языкоглоточного нерва; 2 - ядра моста; 3 - пирамидный тракт; 4 - ядро нижней; оливы](pic/000122.jpg)
Рис. 111. Схема строения ствола мозга. У типичного нейрона ретикулярной формации аксоны направлены как каудально, так и краниально, а дендриты ветвятся по поперечным плоскостям (А). Это навело на мысль, (см., например, [223]) о том, что РФ можно упрощенно представить себе в виде стопки игральных фишек (Б). 1 - ядро языкоглоточного нерва; 2 - ядра моста; 3 - пирамидный тракт; 4 - ядро нижней; оливы
Соответственно Килмер и Мак-Каллок положили в основу своей модели представление о "модуле" - некотором формализованном участке нейропиля, что позволило им избежать одновременно обеих крайних позиций - представления об РФ как о чем-то едином и неплодотворных попыток изучать функцик РФ прямо на нейронном уровне. Следует подчеркнуть, что разбиение РФ на такие модули достаточно произвольно, хотя изменение комбинаций сенсорных модальностей по мере продвижения от головного конца к хвостовому и согласуется с идеей подобной сегментации.
Как афферентный восходящий спино-ретикулярный тракт, так и эфферентный нисходящий пирамидный тракт проходят через сегменты нейропиля (фишки) в перпендикулярном направлении, отдавая коллатерали в эти сегменты. В каждом сегменте имеются нейроны, дендритные поля которых в значительной мере перекрываются. Входные воздействия для каждого конкретного сегмента не специфичны в отношении сенсорной модальности, т. е. на любую клетку РФ оказывает влияние широкий диапазон афферентных стимулов. Выходные аксоны клеток РФ идут как вверх, так и вниз по стволу на весьма большие расстояния, отдавая многочисленные коллатерали. Создается впечатление, что каждый модуль в этом наборе образует разветвленные связи с другими модулями и оказывает весьма сложное выходное воздействие на многие отделы нервной системы.
Перейдем теперь к рассмотрению структуры, приведенной на рис. 112; она состоит из ряда модулей, каждый из которых соединен с несколькими соседними модулями и получает разные входные воздействия, хотя входы соседних модулей могут иметь и много общего. Допустим, что каждый из этих модулей, основываясь на только что полученной информации, может принимать предварительные решения об относительной пригодности того или иного типа поведения. После этого мы можем поставить основной вопрос.
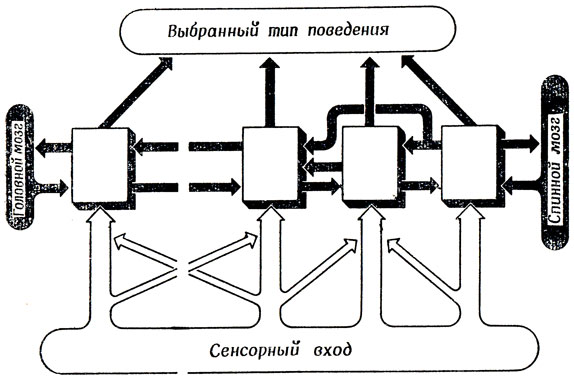
Рис. 112. Развитие идеи о стопке фишек (рис. 111). Теперь мы моделируем РФ как набор модулей. Задача состоит в том, чтобы найти такую конструкцию этой модульной системы, которая давала бы возможность организму выбирать единое согласованное поведение, несмотря на то что все модули получают различную входную сенсорную информацию
Вопрос о согласовании локальных решений. Лак должно быть организовано взаимодействие между модулями, чтобы при наличии разных входных воздействий на разные модули, обусловливающих вначале выбор разных типов поведения, в конечном итоге достигалось некое согласованное решение, определяющее единый тип поведения для системы в целом?
Все дальнейшее следует, по-видимому, рассматривать как важную, хотя и чисто спекулятивную попытку ответить на этот вопрос. Другими словами, воспользовавшись биологическими данными, указавшими нам на важную роль РФ в выборе типа поведения, и анатомическими данными, позволяющими считать схему на рис. 112 адекватной основой для нашей модели, мы теперь забудем о данных нейрофизиологии и используем все что можно, чтобы заполнить блоки на этой схеме и получить конструктивный ответ на вопрос о согласовании локальных решений. Мы увидим, что Килмеру и Мак-Каллоку действительно удалось найти ответ на этот вопрос, но нам придется вновь заняться экспериментированием, чтобы выяснить, соответствует ли принятая ими детализация функций модуля тому, что можно обнаружить в ограниченном сегменте нейропиля настоящей РФ, а затем произвести теоретические исследования, с тем чтобы объяснить, как реальные нейронные сети могут реализовать эти функции модулей, постулированные на основе как теоретических, так и экспериментальных соображений. Поэтому повторим еще раз, что, хотя описываемая ниже схема и дает возможность согласовывать локальные решения, вопрос о том, в какой степени она соответствует процессам, происходящим в стволе головного мозга реально существующих животных, остается открытым.
Для простоты изложения предположим, что мы хотим построить блок выбора, подобный схеме 112, всего из четырех модулей, так что выбирать приходится всего ,из четырех типов поведения. Опишем состояние каждого модуля вектором p=(p1, p2, p3, p4), где каждое pj≥0, а p1+p2+p3+p4=1 и каждое pj(j=1, 2, 3, 4) описывает вес (например, адекватность), который этот модуль в настоящий момент приписывает j-му типу поведения. Будем считать, что локальные решения согласованы, если более 50% модулей приписывают одному и тому же типу поведения вес, превышающий 1/2, и именно в такое поведение и оказывается вовлеченной система. К сожалению, Килмер и Мак-Каллок не уделили никакого внимания ни поискам правдоподобной нейронной системы обнаружения самого факта достижения согласия, ни тому, как должны быть устроены системы управления движением, с тем чтобы они могли реагировать на указания о выбранном типе поведения. В качестве первого шага к решению первой из этих проблем отметим, что, как показывает машинное моделирование, всякий раз, когда процесс сходится, на выходе очень скоро возникает ситуация, при которой не менее 70% модулей приписывают доминирующему типу поведения вес не ниже 0,65. В связи с этим для каждого типа поведения можно предусмотреть свой собственный блок порогового сравнения, срабатывающий лишь тогда, когда входное воздействие достигнет по крайней мере 50% своего максимума, и не опускается ниже этого уровня на протяжении некоторого времени. Это гарантирует, что в любой момент времени проявит активность лишь один регулятор типа поведения и, следовательно, организм будет отрабатывать поведение только одного типа. Такой детектор согласования использовался в процессе машинного моделирования, однако вопрос о том, в какой форме эта операция может реализовываться в нервной системе, остается нерешенным.
Стратегию, воплощенную в схеме Килмера и Мак-Каллока, можно сравнить с обменом мнениями между врачами, собравшимися на консилиум, чтобы поставить диагноз. Каждый из участвующих в консилиуме врачей - специалист в определенной области, но достаточно хорошо знаком и с медициной в целом. Кардиолог может основывать свой диагноз главным образом на характере электрокардиограммы, но не может не обратить внимания на бледность больного и на его одышку. Осматривая пациента впервые, врачи (наши модули) "глобально независимы" и каждый самостоятельно ставит первоначальный диагноз. Но затем начинается дискуссия с целью выработать единое мнение. В процессе этого обмена могут возникать и ситуации "локальной автономии" мнений; если один из врачей неожиданно почувствует уверенность в том, что диагноз должен быть совершенно иной, нежели тот, который он отстаивал ранее, то в течение определенного периода времени остальные врачи будут уделять особое внимание причинам, побудившим первого врача изменить свое мнение.
Основываясь на подобных соображениях, Килмер, Мак-Каллок и Блум [137] создали машинную модель, названную ими S-ретической. Это была модульная система, предназначенная для выбора типа поведения и работавшая по следующей схеме:
1. Если входное воздействие на модуль изменяется очень сильно, то новое значение p определяется практически только характером нового сенсорного входа. Это позволяет обеспечить глобальную автономию модулей при изменениях входа системы в целом.
2. Если входное воздействие на модуль изменяется не oслишком сильно, то новое р определяется нормированным взвешенным средним старого p и p других модулей, связанных с рассматриваемым. При этом локальная автономия достигается путем приписывания большего веса тем p, которые претерпевают внезапные изменения, а избыточность потенциальных команд получается в результате приписывания большего веса тем p, которые содержат больше селективной информации.
Как уже отмечалось, мы (произвольно) считаем, что решение согласовано, когда большинство модулей приписывают одному и тому же типу поведения вес, больший 0,5. Поэтому суммарный эффект работы схемы можно рассматривать как изначальное обеспечение автономии модулей в момент появления нового входа, позволяющее акцентировать внимание на даваемых каждым модулем оценках того, каким должно быть новое поведение, с последующим итеративным увязыванием индивидуальных решений в одно глобальное, согласованное. Моделирование на вычислительной машине показывает, что в S-ретической схеме (или по крайней мере в конкретной рассматривавшейся системе взаимосвязей между модулями) искомое согласование достигается не более чем за 25 итераций и после того, как оно будет достигнуто, сохраняется до появления нового входа. Если входные воздействия ясно указывают на адекватность лишь одного типа поведения, согласование достигается весьма быстро. Но если предпочтение одного типа поведения не вполне ясно выражено, то начальные условия и характеристики системы оказывают сильное влияние на окончательное решение.
В рамках поведения одного и того же типа возможны многие различные действия. Например, если кошка решает спасаться бегством, то она может утащить мышь с собой или бросить ее может вскарабкаться на дерево или обежать его, перепрыгнуть через ручей или переплыть его. Килмер и Мак-Ларди [136] считают, что функцию выбора действий в пределах поведения данного типа выполняет одна из областей гиппокампа (остальные его части заняты, по крайней мере в период формирования поведения, по-видимому, консолидацией долговременной памяти). Мы не собираемся рассматривать здесь модель, предложенную этими авторами, поскольку она остается пока весьма спорной (ср., например, с другой моделью гиппокампа, принадлежащей Олдзу [194]), но, возможно, читатель захочет поразмыслить над. тем, почему иерархическая структура системы выбора поведения и конкретного действия в рамках выбранного типа поведения может быть в определенном смысле "лучше" (независимо от вида конкретной структурной схемы, реализующей эти функции) другой, в которой последовательные действия выбираются: сразу.
Возвращаясь к выбору поведения, отметим, что утверждение "РФ управляет выбором типа поведения" не следует понимать - в том смысле, что РФ может нейтрализовать любое воздействие коры головного мозга. Напротив, кора может играть здесь важнейшую роль, устанавливая относительную важность различных, комбинаций входных стимулов в выбранном РФ типе поведения.. Например, один из важных параметров изменения типа поведения- это состояние готовности. Обычно животное постепенно переходит от глубокого сна к чуткому, затем просыпается и, наконец, оказывается в состоянии полной готовности. Неожиданный звук может сразу привести животное в состояние готовности, и, по-видимому, можно считать вполне установленным, что это происходит в результате передачи возбуждения по нервным путям, распространяющимся диффузно в кору головного мозга: из области РФ, получившей в связи с этим название активирующей ретикулярной системы. Однако при многократном повторении одного и того же звука РФ может перестать активизировать животное, и в этом случае мы говорим, что животное привыкло к данному стимулу. Такое привыкание может быть достаточно сложным. Спящая женщина может совершенно не реагировать на раскаты грома, но просыпается, как только заплачет ее ребенок; матрос может мирно спать под грохот судового двигателя и мгновенно просыпаться, если эти привычные звуки вдруг изменятся. Состояние готовности может быть вызвано и отсутствием привычного стимула. Так, Прибрам [204] рассказывает, что жители Нью-Йорка, привыкшие к грохоту поезда надземной железной дороги, проходившего в 3 часа ночи, в течение долгого времени после того, как эту дорогу снесли, просыпались ровно в 3 часа ночи, пока не привыкли к необычной тишине! Профессор Московского государственного университета Е. Н. Соколов поставил опыт, показывающий, что в мозгу, по-видимому, формируется "внутренняя модель" стимула, к которому мы привыкаем, и что изменение достаточного числа произвольных параметров этой модели может привести к исчезновению привычки. В опыте Соколова испытуемый многократно подвергался звуковому воздействию с фиксированной частотой и продолжительностью. При этом по мере повторения ориентировочный рефлекс, измерявшийся "вызванной реакцией" (одна из грубых мер уровня электрического возбуждения коры), становился все более и более слабым. Напрашивалось предположение, что РФ просто научилась игнорировать слуховые воздействия этой частоты. Однако Соколов [236] опроверг такое объяснение, продемонстрировав, что привычка к этой частоте исчезает, если повысить или даже понизить интенсивность стимула. Более того, если после того, как к стимулу заданной частоты и амплитуды выработалась привычка, резко уменьшать его продолжительность, то в течение этого "недостающего времени" будет наблюдаться вызванная реакция. Таким образом, в организме закодированы и частота, и амплитуда, и продолжительность стимула, и привыкание объясняется, по-видимому, не утомлением или отключением какой-то ограниченной группы периферических рецепторов или детекторов признаков, а скорее централизованным формированием "эталона" или набора параметров, используемых при эфферентном управлении соответствующими участками сенсорных путей, что позволяет использовать в проводимых в РФ операциях только информацию о рассогласованиях с эталоном.
Впрочем, здесь необходимо отметить, что при удалении коры погасить ориентировочный рефлекс удается лишь в самых необыкновенных случаях. Так, хотя этот рефлекс и управляется РФ, кора головного мозга может оказывать тормозное воздействие на оценку сенсорного входа. Поэтому модель Килмера - Мак-Каллока объясняет лишь то, каким образом РФ могла бы .выбирать тип поведения при отсутствии воздействий со стороны коры. Мы надеемся, что нам удастся лучше разобраться в поведении человека, если мы попытаемся проследить за постепенным совершенствованием подобных примитивных операций в процессе эволюции, вместо того чтобы стараться понять это поведение сразу во всей его сложности. Аналогичным образом мы пытаемся разобраться в организации зрительного восприятия, выясняя, в какой степени зрительную кору можно рассматривать как результат эволюции, приведшей к совершенствованию системы примитивной визиомоторной координации, осуществлявшейся в тектуме. Как мы видим и в том и в другом случае, коре приходится перерабатывать много информации, чтобы помочь структурам более низкого уровня решать, что же должно быть в центре нашего внимания, и приходится признать, что наши представления об этих процессах находятся еще на самом начальном этапе.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'