2.4. Предварительная обработка зрительной информации у кошки и лягушки
Эттнив [18] отмечает, что, располагая некоторой априорной информацией о какой-либо совокупности "сообщений", можно использовать эту информацию для экономного описания этого множества, недоступного в противном случае. Например, если бы любые сочетания черных и белых точек образовывали одинаково информативные для зрительного восприятия картинки в повседневной жизни, то не существовало бы более экономичного способа представления увиденного, чем описание интенсивности освещения каждой точки мозаичного зрительного поля. Однако, как свидетельствует успех карикатур, большой объем зрительной информации можно передать с помощью нескольких контурных линий. Более того, эти контуры обычно можно очертить с помощью относительно небольшого числа штрихов. Торчащие в разные стороны волоски на спине кошки, нарушающие ее плавный изгиб, несущественны для нашего узнавания очертаний кошки, хотя само распознавание "текстуры" шкурки и может дополнить наше восприятие. Основываясь на этих соображениях, Эттнив высказал предположение, что в распознавании формы наиболее важную роль играют те точки, в которых контурные линии меняют свое направление или обрываются. Например, он построил рис. 19, найдя на контуре изображения спящей кошки 38 точек максимальной кривизны и соединив соответствующие точки прямыми.

Рис. 19. Спящая кошка Эттнива. Этот рисунок, на котором легко узнать кошку, получен в результате замены естественных контуров прямыми, соединяющими точки максимальной кривизны
Однако он не рассматривал вопрос о том, как выяснить, какие пары точек нужно соединять между собой. Простое решение этой проблемы показано на рис. 20: надо, по-видимому, не просто отмечать точки максимальной кривизны (А), ибо это может привести к неправильным соединениям (Б), но дополнить их данными об углах (В), что позволит получить правильное решение (Г).
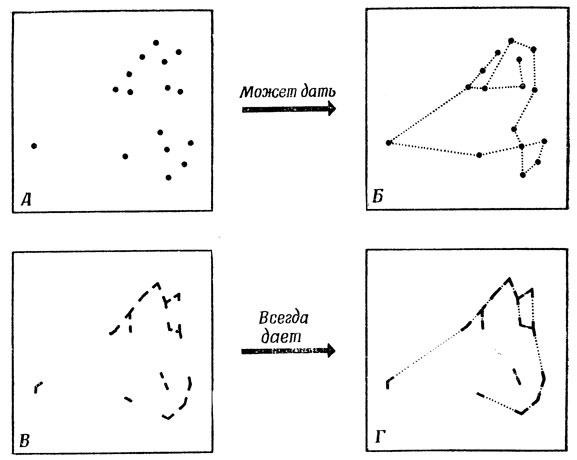
Рис. 20. О пользе информации об углах
Все это позволяет считать, что полезная схема предварительной обработки зрительных стимулов может слагаться из двух этапов: 1) сведение картины распределения света и тени к выделению основных контуров; 2) переход от контурных линий к измерению углов в точках изменения направления контура.
Барлоу [20] подошел к проблеме предварительной обработки зрительной информации на основе более строгих нейрофизиологических соображений. Считая, что рецепторы играют роль датчиков, преобразующих энергию внешней среды, скажем, в частоту возбуждения аксонов, он задал себе вопрос: а как последующие уровни обработки (рис. 21) могли бы, используя закономерности, характерные для обычной среды, регистрировать входные сигналы таким образом, чтобы "минимизировать транспорт импульсов по нервным путям", т. е. обеспечить ситуацию, при которой активность нейрона отклоняется от уровня покоя только в том случае, когда необходимо сообщить животному о некотором потенциально важном для него свойстве внешней среды? Как отметил Барлоу [21], работы Эттнива наводят на мысль, что подходящий набор устройств предварительной обработки выделяет точки максимальной кривизны, а наше рассмотрение проблемы соединения таких точек подсказывает, что для соответствующих областей потребуются и детекторы угла.
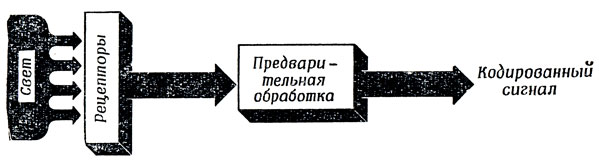
Рис. 21. Схема организации предварительной обработки зрительного образа. Рецепторы преобразуют распределение интенсивности света в градуальные изменения мембранного потенциала. Следующие за ними слои нейронов обеспечивают предварительную обработку этих потенциалов и кодируют то, что видит глаз, в форме пространственно-временной структуры спайков, поступающей в мозг
Для того чтобы конкретизировать эти общие соображения, полезно сопоставить зрительные системы кошки и лягушки. У всех позвоночных, будь то лягушка, кошка или человек, свет попадает в глаз через хрусталик и направляется к задней стенке глаза; пройдя через несколько слоев прозрачных клеток сетчатки, он наконец попадает на колбочки и палочки (рис. 22). При достаточной интенсивности света в палочках и колбочках возникает генераторный потенциал, вызывающий импульсы в аксонах ганглиозных клеток через посредство вставочных нейронов, аксоны которых образуют синапсы с дендритами этих клеток. Зрительные волокна - это аксоны ганглиозных клеток, которые, пересекая сетчатку, собираются в области, называемой слепым пятном; здесь они образуют зрительный нерв, который направляется в мозг.
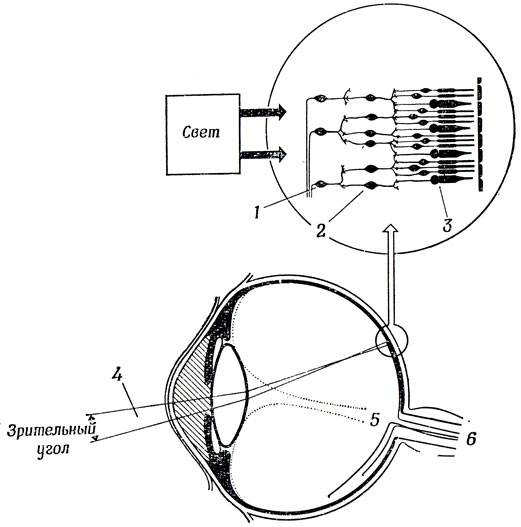
Рис. 22. Схема, показывающая, как хрусталик осуществляет фокусировку' света на сетчатку. Увеличенное изображение фрагмента сетчатки (вверху) показывает, как пучок света про ходит к задней стенке сетчатки, возбуждая палочки и колбочки (3), а те в свою очередь возбуждают вставочные нейроны (2), управляющие одной из ганглиозных клеток (1). Проследив ход световых лучей в обратном направлении от рецепторов, воздействующих на определенную ганглиозную клетку, и через хрусталик, можно найти участок внешней среды-рецептивное поле данной клетки (4), освещенность которого влияет на характер импульсации этой клетки. Аксоны ганглиозных клеток проходят через сетчатку и, собираясь вместе в слепом пятне (5), образуют зрительный нерв (6), идущий в мозг
Поскольку, как мы более подробно покажем ниже, выходные сигналы ганглиозных клеток резко отличаются от кодированных сигналов об интенсивности светового потока, генерируемых рецепторами, можно утверждать, что в сетчатке происходит предварительная обработка зрительных стимулов. Напомним главные моменты анатомии зрительной системы млекопитающих (разд. 2.3), например человека и кошки. Прошедшие предварительную обработку сообщения посылаются в латеральное коленчатое тело (рис. 23, А) - "промежуточную станцию" зрительного тракта в таламусе, - а после дальнейшей обработки в различные клетки коры, где происходит распознавание образов, тогда как сигналы, идущие по дополнительным (но более древним с эволюционной точки зрения) каналам используются механизмами ориентации (например, поворота головы и туловища), расположенными в верхнем двухолмии (разд. 5.4).
У лягушки (рис. 23, Б) зрительные нервы идут от сетчатки назад, в основном к зрительному бугру. Стрелки показывают, как зрительные стимулы проецируются на зрительный бугор, который у лягушек в отличие от млекопитающих играет главную роль в распознавании образов ("Как связать единое целое?") и в ориентации ("Где находится?"). Анатомическое сходство верхнего двухолмия млекопитающих со зрительным бугром лягушки, возможно, свидетельствует о некотором сходстве функций этих структур, но, как станет ясно ниже, это не указывает на их полную функциональную идентичность.
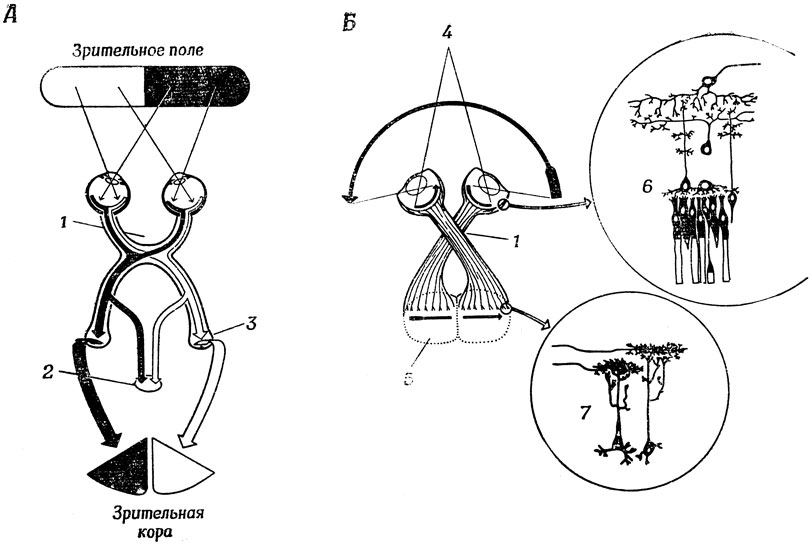
Рис. 23. Сравнительный анализ зрительной системы человека (А) и лягушки (Б). У человека большинство сообщений проходит через ядро латерального коленчатого тела таламуса в кору головного мозга, и лишь немногие из них попадают в верхнее двухолмие. У лягушки же преобладают связи со зрительным бугром (аналог верхнего двухолмия млекопитающих). Отметим также отчетливое разделение зрительного поля человека да две части, совершенно отсутствующее у лягушки. 1 - хиазма; 2 - верхнее двухолмие; 3 - латеральное коленчатое тело; 4 - глазные яблоки; 5 - зрительный бугор; 6 - сетчатка лягушки; 7 - нейроны зрительного бугра лягушки
Рецептивное поле ганглиозной клетки (у кошки или у лягушки) - это область зрительного поля, раздражение которой может влиять на активность данной ганглиозной клетки. Как показано на рис. 22, рецептивное поле представляет собой пространственный конус, в пределах которого свет попадает на те колбочки и палочки, которые возбуждают вставочный нейрон, управляющий данной ганглиозной клеткой. Тем самым зрительный угол рецептивного поля определяется величиной угла при вершине этого конуса.
Каков характер световой картины, на которую реагирует ганглиозная клетка? Другими словами, какая картина вызывает максимальное возбуждение этой клетки? Как это ни удивительно, но, хотя строение сетчатки у всех позвоночных в основном одинаково и соответствует схеме на рис. 22, функции ганглиозных клеток сильно различаются у разных видов. Например, Леттвин, Матурана, Мак-Каллок и Питтс [150] выяснили, что большая часть ганглиозных клеток лягушек принадлежит к одному из четырех типов, например к "детекторам движения" или к "детекторам закругленного края", а у кошек ганглиозных клеток ни одного из этих типов нет вовсе, а есть "устройства для усиления контраста". Надо сказать, что это вполне согласуется с нашим принципом деенаправленного восприятия. У лягушки, в поведении которой зрительная информация играет незначительную роль, если не считать схватывания "вихляющихся" объектов и бегства от "врагов", сетчатка "отбрасывает" большинство аспектов зрительных образов, не связанных с этими чертами, тогда как кошка с ее гораздо более сложным поведением (например, пристальное наблюдение за мышиной норой и выбор момента для прыжка только после того, как мышь достаточно далеко высунется из норы) не могла бы существовать, если бы ее сетчатка была так сильно специализирована. Проанализируем эксперименты по предварительной обработке зрительной информации более тщательно.
Обычно лягушка не двигает глазами, за исключением тех случаев, когда ей необходимо компенсировать движения головы и туловища (например, если она сидит на качающемся листе кувшинки) для стабилизации изображения на сетчатке. Поэтому, хотя лягушки обнаруживают свою добычу исключительно благодаря зрению, они не следят за добычей и не сканируют зрительное поле в поисках объектов, представляющих для них интерес. Они охотятся лишь за движущимися насекомыми, а неподвижные объекты не привлекают их внимания. Появление большого движущегося объекта вызывает у них реакцию бегства в наиболее затемненную часть окружающего пространства, а так как лягушка одинаково хорошо чувствует себя и в воде, и на суше, то выбор пункта конечного назначения побега не представляется существенным. Для лягушек неподвижный предмет с поведенческой точки зрения лишен смысла. По-видимому, они узнают добычу и схватывают ее, выбирая среди других объектов окружающей среды на основании ряда признаков, таких, как движение, определенные размеры, контрастность, а может быть, также и окраска. Поэтому на дергающуюся лапку другой лягушки, которая решила почесать себе спинку, они реагируют как на летящую муху, но они никогда не схватывают мертвую муху, даже погибая от голода. Способность лягушки распознавать свою добычу и схватывать ее не изменяется при изменении внешней обстановки, например освещенности. В их половом поведении главную роль играют слух, осязание и обоняние.
Леттвин, Матурана, Мак-Каллок и Питтс [150] исследовали зрительную систему лягушки с целью выявить те ее свойства, которые помогают лягушке узнавать свою добычу и врагов. В отличие от Хартлайна [106], разбившего все ганглиозные клетки на три класса в зависимости от их реакции на маленькое световое пятно в соответствующем рецептивном поле, Леттвин и др. обратили внимание на то, что световое пятно не служит таким естественным стимулом для лягушки, как муха или червяк (если только она не питается светлячками), и постарались установить более "натуральные" функции ганглиозных клеток, изучая их реакцию на естественные объекты. Они исходили из того, что поведение лягушки требует, по-видимому, существования каких-то функциональных инвариантов в активности компонентов ее зрительной системы, с тем чтобы сетчатка в результате обработки зрительного образа могла выделять его важные черты, например присутствие добычи или опасности.
Здесь мы расскажем о тех зрительных коррелятах активности, передаваемой ганглиозными клетками в зрительный бугор, которые им удалось обнаружить. Позже в разд. 7.2 мы вернемся к этим данным для того, чтобы построить модель преобразования активности в зрительном бугре лягушки в действие, т. е;. на основе теории "что глаз лягушки говорит ее мозгу" мы построим гипотезу о том, "что глаз лягушки говорит самой лягушке". Леттвин и др. работали на обычной американской лягушке Rana pipiens. Лягушку помещали в экспериментальную установку так, чтобы один ее глаз оказался в центре полушария (рис. 24) диаметром 35,5 см (при этом 1° зрительного угла соответствовала дуга длиной 3 мм), составлявшего зрительное поле. В голову лягушке вживляли электрод, который регистрировал активность либо единичной ганглиозной клетки (для этого кончик электрода помещали на аксон этой клетки в зрительном нерве), либо одной из клеток зрительного бугра. Алюминиевое полушарие охватывало примерно две трети зрительного поля одного глаза лягушки. Расположив животное соответствующим образом, можно получить возможность перекрыть любую часть зрительного поля и полностью контролировать рецептивное поле изучаемых клеток. Стимулирующие объекты перемещали по внутренней поверхности полушария с помощью магнита, двигавшегося по внешней поверхности. В опытах использовались самые разнообразные объекты, например, темные диски, темные полоски и темные квадраты.
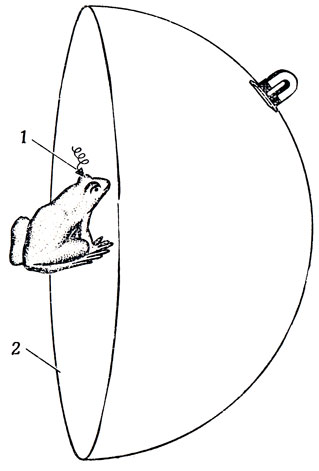
Рис. 24. Магнит на внешней поверхности алюминиевого полушария (2) позволяет экспериментатору перемещать зрительные стимулы по внутренней поверхности полусферы. Реакцию лягушки на эти стимулы можно снимать с помощью микроэлектродов (1), вживленных в зрительный нерв или зрительный бугор
Оказалось, что по своей реакции ганглиозные клетки распадаются на четыре типа (возможно, имеется и пятый тип, но о нем мы здесь говорить не будем). Каждая ганглиозная клетка принадлежит только к одному из этих типов, а клетки каждого типа равномерно распределены по сетчатке. На любом небольшом участке сетчатки можно найти клетки всех четырех типов в соотношении, соответствующем их общему соотношению в сетчатке в целом. Так, клетки типов I и II встречаются в 30 раз oчаще клеток типов III и IV.
Тип I - детекторы границы (угловые размеры рецептивного поля 2-4°). Реагируют на границу между светлыми и темными участками, попадающую в рецептивное поле, независимо от ориентации границы, но при условии, что она достаточно резкая. При этом скорее всего измеряется резкость границы, а не степень контраста между соседними участками. Реакция усиливается, если границу сдвинуть, и практически не изменяется,, если менять освещенность в очень широких пределах. Если в рецептивное поле такая граница не попала, то изменение освещения поля, каким бы резким оно ни было, не вызовет никакой реакции. Еще одно свойство клеток типа I заключается в том,, что если границу внести в рецепторное поле в полной темноте, а затем включить свет, то продолжительная реакция возникает лишь с некоторым запозданием.
Тип II - детекторы движущегося закругленного края, или детекторы насекомых (угловые размеры рецептивного поля 3-5°). Реагируют также лишь на резкую границу между светлым и темным, но только при дополнительных условиях: граница должна быть изогнута и притом так, чтобы более темная область была выпуклой и чтобы граница двигалась. Как и в первом случае, реакция клетки не меняется от изменения освещенности поля в широких пределах, соответствующих, грубо говоря, изменениям освещения от сумеречного до полуденного. В одном интересном опыте в рецептивное поле помещали большую цветную фотографию естественного места обитания лягушек с цветами и травой. Покачивание этой фотографии в рецептивном, поле волокна клетки типа II не вызвало никакой реакции. Когда на фоне этой картинки перемещали предмет, похожий на муху, то лягушка бурно реагировала, если же этот предмет покачивали вместе с фотографией, то это ни к чему не приводило.
Тип III - детекторы движущейся границы (угловые размеры рецептивного поля 7-11°). Эти клетки в широком диапазоне изменения освещенности одинаково реагируют на один и тот же силуэт, движущийся с одной и той же скоростью на одинаковом фоне. Их реакция непродолжительна, и они возбуждаются лишь в том случае, если граница изменяется или перемещается. Реакция выражена тем сильнее (частота ее тем выше), чем резче граница и чем быстрее она движется.
Тип IV - детекторы затемнения (угловые размеры рецептивного поля 15°). Реагируют на любое снижение освещенности поля в целом с учетом расстояния до центра этого поля. Границы не оказывают на реакцию этих клеток никакого влияния. Одинаковое относительное снижение вызывает одинаковую реакцию, более или менее не зависящую от исходного уровня освещенности. Продолжительность реакции увеличивается, если какой-либо большой темный предмет останавливается в пределах поля.
Аксоны детекторов каждой группы оканчиваются в разных слоях зрительного бугра. Каждый из этих четырех слоев окончаний в зрительном бугре образует "непрерывную" карту сетчатки применительно к операциям, выполняемым соответствующими ганглиозными клетками. Эти четыре слоя регистрируют приходящую информацию, причем точки, относящиеся к различным слоям и лежащие в зрительном бугре друг над другом, соответствуют одному и тому же небольшому участку сетчатки (рис. 25).
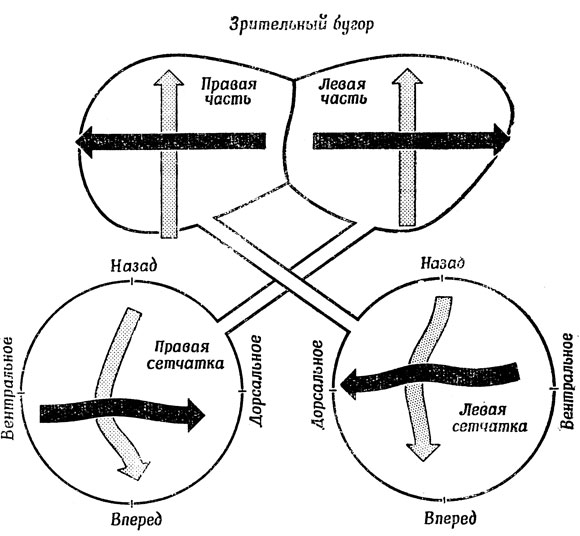
Рис. 25. Ретино-тектальная проекция у взрослой жабы, показывающая, как некое направление движения, регистрируемое сетчаткой, отображается в соответствующее направление в зрительном бугре
Таким образом, функция сетчатки лягушки состоит не в том, чтобы передавать информацию о поточечном распределении света и тени в рассматриваемом образе; ее главная функция - анализ этого образа в каждой точке в поисках четырех различных качественных признаков (границ, движущегося закругленного края, движущейся границы и локального затемнения) и передача этой информации в зрительный бугор, где она распределяется между четырьмя слоями нервных окончаний.
Это означает ([150], стр. 1950), что "глаз общается с мозгом на языке, который уже подвергся некоторой организации и переработке, а не просто передает более или менее точную копию распределения света на рецепторах". Более того, используемое при этом кодирование помогает лягушке находить пищу и избегать врагов, т. е. распознавать универсали "добыча" и "враг". Эту мысль можно найти в заключительном абзаце (там же, стр. 1951) статьи Леттвина и др., который я позволю себе процитировать:
"Таким образом, эти операции гораздо больше напоминают восприятие, чем ощущение, если в настоящее время имеет смысл проводить между ними различие. Иными словами, эти операции удобнее всего описывать на языке сложных абстракций зрительных образов. Нам казалось, например, весьма соблазнительным назвать детектор закругленного края "детектором насекомых". Соответствующее волокно (типа II) реагирует наиболее интенсивно, когда в его рецептивное поле входит темный объект меньший, чем это поле, останавливается, а затем начинает метаться из стороны в сторону. На реакцию волокна не влияет ни изменение освещения, не перемещение фона (например, картинки с изображением травы и цветов). Если же в рецептивное поле попадает только фон, движущийся или неподвижный, то никакой реакции не наблюдается. Так можно ли лучше описать систему, предназначенную для обнаружения насекомых, находящихся в пределах досягаемости?"
Перейдя от ганглиозных клеток к клеткам зрительного бугра, Леттвин и его сотрудники обнаружили и там клетки нескольких разных типов. Им не удалось сколько-нибудь удовлетворительно установить функции клеток разных типов, однако они выделили нейроны двух крайних типов, названные "нейронами новизны" и "нейронами постоянства". Функция первых состоит, по-видимому, в том, чтобы обнаруживать новое и подмечать зрительные события, а вторые обеспечивают непрерывное слежение за интересным объектом, попавшим в поле зрения. Нейроны по крайней мере одного из этих двух типов посылают аксоны обратно в сетчатку и оказывают существенное влияние на активность ганглиозных клеток.
Сравним теперь систему предварительной обработки зрительной информации у лягушки с аналогичной системой у кошки. При этом мы воспользуемся работой Хьюбела и Визеля, посвященной зрительной системе кошки. У кошки возбуждение от рецепторов передается различным вставочным нейронам. В свою очередь биполярные вставочные нейроны возбуждают ганглиозные клетки сетчатки, а тормозное воздействие, по-видимому, осуществляют вставочные нейроны другого типа, называемые горизонтальными клетками. Таким образом, на каждую ганглиозную клетку воздействует целая система рецепторных клеток. Как установил Каффлер, у кошки рецептивные поля ганглиозных клеток сетчатки в первом приближении имеют форму диска с двумя различными областями (рис. 26). Колбочки и палочки, воздействующие через посредство вставочных нейронов на центральную область, обычно оказывают возбудительное влияние, т. е. повышение интенсивности света, падающего на колбочки или палочки, связанные с этой центральной зоной, повышает частоту импульсов этой клетки; напротив, связи между рецепторами и периферической областью поля организованы так, что усиление интенсивности падающего на эти рецепторы света вызывает снижение активности ганглиозной клетки. При этом эффективность синапсов по всему рецептивному полю такова, что если равномерно осветить все это поле, то возбуждение, создаваемое центральной областью, незначительно превысит торможение, создаваемое периферической областью, и в результате в клетке возникнет слабая спонтанная импульсация, несущая информацию об интенсивности такого равномерного освещения*. Имеются также и клетки, у которых центральная область рецептивного поля является тормозной, а периферическая - возбудительной.
* (Между прочим, Хьюбел и Визель обнаружили, что у обезьян наряду с колбочками и палочками, чувствительными к разным цветам и максимально реагирующими одни на красный, другие на зеленый, а третьи на синий цвет, имеются еще и ганглиозные клетки, способные обнаруживать границы между участками разного цвета (например, зеленый кружок на красном поле). Однако здесь мы будем рассматривать лишь монохроматический свет.)
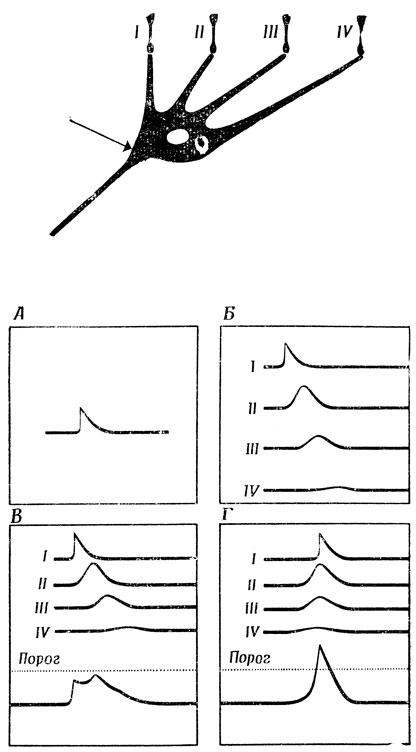
Рис. 26. Реакции двух типов ганглиозных клеток кошки. Верхняя схема изображает клетку с возбудительной центральной областью (on-центром) и тормозной периферией (off-периферией), а нижняя - с тормозной центральной областью и возбудительной периферией. Справа показаны типичные записи импульсов, которые можно получить при экспериментах на таких клетках. Первый участок соответствует низкочастотному спонтанному возбуждению такой клетки в результате продолжительного пребывания в темноте, следующий - записи, полученной при появлении в указанной области рецептивного поля небольшого источника света. Например, для клетки с on-центром появление источника света в центральной области поля резко повышает частоту импульсов, а появление этого же источника на периферии столь же резко снижает эту частоту. Наконец, правый участок дает представление о преходящих изменениях частоты импульсов, возникающих сразу после выключения света и предшествующих возвращению к норме
Весьма упрощенный пример ясно покажет, как свойства нервной ткани, описанные в разд. 2.2, дают возможность простому нейрону за счет одной геометрии дендритного ветвления вычислять некоторые полезные функции. Прежде всего мы построим модель нейрона, который может обнаруживать последовательность входных сигналов с заданными временными соотношениями (рис. 27), а затем изучим клетку типа ганглиозной клетки кошки, выполняющей роль "детектора контраста". Сначала рассмотрим нейрон с четырьмя дендритами (ср. [209], стр. 90); каждый дендрит образует по одному синапсу со зрительным рецептором, причем синапсы I-IV расположены на все возрастающем расстоянии от аксонного холмика этой клетки (рис. 27).
Предположим, что каждый рецептор реагирует на проходящее над его поверхностью световое пятно возбуждением генераторного потенциала, создающего в постсинаптической мембране - деполяризацию, динамика которой показана на рис. 27,А и которая накладывается на потенциал покоя. Эта деполяризация распространяется пассивно, и чем на большее расстояние она передается, тем позже возникает пик и тем он меньше (рис. 27,Б). Эффект возбуждения синапса IV проявляется в аксонном холмике позже и в меньшей степени, чем возбуждение- синапсов I, II или III. Рассмотрим теперь, к чему это приводит. Если все входные возбуждения приходят в синапсы I-IV одновременно, то их воздействие на аксонный холмик можно приближенно представить суммой индивидуальных эффектов - (рис. 27,В). Пусть этот суммарный эффект меньше порога, необходимого для того, чтобы вызвать появление спайка. Создается впечатление, что такая клетка вообще бесполезна поскольку она не реагирует на то, что на первый взгляд кажется максимальным стимулированием, а именно - на одновременное раздражение всех ее четырех входов. Однако, если входной сигнал попадает на вход IV раньше, чем на вход III, а на вход III раньше, чем на вход II, и т. д. и при этом интервалы между входными сигналами будут такими, что пики всех четырех входных воздействий придут в аксонный холмик одновременно, их суммарный эффект может оказаться существенно больше порога (рис. 27,Г). А это значит, что наша клетка, несмотря на ее крайнюю простоту, может распознавать направление движения относительно ее входов.
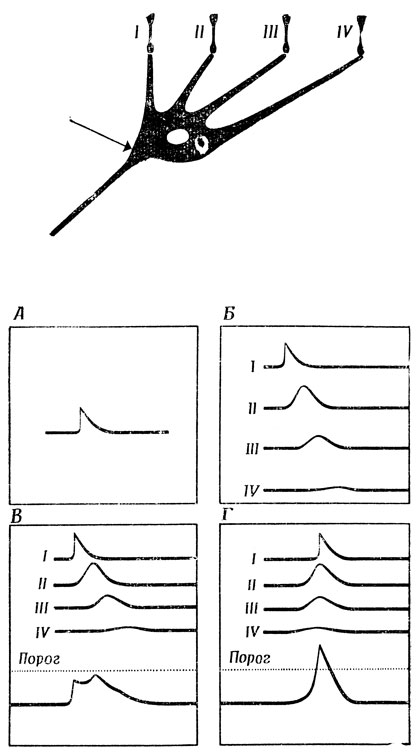
Рис. 27. Модель нейрона, способного выполнять функции детектора движения. Изменение потенциала (А), происходящее в синапсе, постепенно затухает, а его пик запаздывает тем больше, чем больше расстояние ему нужно пройти (Б). Поэтому если все четыре стимула приходят на синапсы I-IV одновременно, то их суммарный эффект в аксонном холмике (стрелка) не достигает порогового (В). Но если эти синапсы возбуждаются в последовательности IV-III-II-I, так что их пики достигают аксонного холмика одновременно, то суммарный эффект оказывается выше порога (Г)
Если представить себе теперь, что к синапсам подходят аксоны фоторецепторов, которые реагируют на пересечение их рецептивного поля светящейся точкой возбуждением генераторного потенциала, то наша ганглиозная клетка будет реагировать лишь в том случае, если эта точка движется справа налево и если скорость этого движения не выходит за известные пределы. Эта клетка не будет реагировать на неподвижный объект или на объект, движущийся слева направо. Таким образом, геометрия клетки может иметь огромное значение для ее функционирования. В рассмотренном примере предпочтение, отдаваемое определенному направлению движения, достигается за счет асимметрии расположения дендритов на теле клетки. Здесь открывается целая область исследований: изучение взаимосвязи между функциями и формой нейронов (обзор работ по этому вопросу см. [209]). И если вспомнить, что на многих нейронах в мозгу человека образуется до 10 000 синапсов, то потенциальное разнообразие функций таких нейронов может показаться нам в зависимости от настроения либо весьма вдохновляющим, либо обескураживающим. Как сказал об этом Баллок, "нейроны - как люди". Социология иногда может быть источником удачных метафор для теории мозга и наоборот.
Рассматривая зрительный мир животного, можно заметить, что наиболее важная зрительная информация связана с изменениями, как пространственными, так и временными. (Вспомните в связи с этим успех дружеских шаржей или о том, как привлекают наше внимание движущиеся предметы.) Соответственно, многие процессы "предварительной обработки" на первых уровнях сенсорной системы направлены как раз на усиление пространственных и временных контрастов, реагируя наиболее сильно на изменения, происходящие в рецептивном поле во времени, или на пространственные изменения в зрительном поле. Познакомившись выше с детектором движущегося светового пятна, рассмотрим теперь детектор пространственных изменений, аналогичный по своим свойствам одной из ганглиозных клеток сетчатки кошки, о которой мы уже упоминали.
Вообще говоря, нам не важно, чтобы освещенность точек А, Б, В и Г была в точности такая, как показано на рис. 28; важно лишь, что при переходе от точки В к точке Г освещение резко меняется. Приведем теперь еще один пример "нейрональной конструкции", используя на этот раз не только возбудительные, но и тормозные синапсы (чтобы приблизиться к ганглиозным клеткам кошки с on-центром и off-периферией), ибо одна из возможностей получить информацию о границе связана с клеткой, соединенной с клетками, расположенными по всей площади круга, причем клетки, расположенные в центре, являются возбудительными, а те, что находятся на периферии, - тормозными (рис. 29). Если обозначить через r0 уровень спонтанной активности клетки, получающей сигнал фоторецепторов в случае равномерно освещенного поля, то при движении рецептивного поля по поверхности с границей так, как это показано на рис. 30, А, изменение реакции клетки во времени дает кривую, представленную на рис. 30, Б. Таким образом, активность клетки отклоняется от r0 в областях, близких к границе.
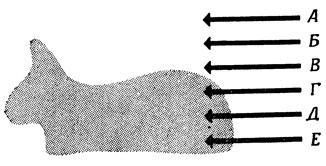
Рис. 28. Важна не абсолютная интенсивность света в точках А, Б, В, Г, Д и Е, а то, что она меняется при переходе от В к Г
Переходя теперь от сетчатки кошки к ее зрительной коре, следует сказать, что Хьюбел и Визель обнаружили там как "простые" клетки коры (которые реагируют на линии, имеющие специфическую ориентацию и расположенные в специфическом месте), так и сложные клетки, реагирующие на линии заданной ориентации, но расположенные в любых участках зрительного поля. Схемы соединений, которые, как они полагают, дают возможность получить поведение, характерное для простых клеток коры, показаны на рис. 31. По-видимому, реакция сложных клеток коры в свою очередь может получить объяснение с помощью относительно простой схемы соединений с соответствующим набором простых клеток коры. Хьюбел и Визель [120] обнаружили еще и "сверхсложные" клетки в полях 18 и 19 (примыкающих к первичной зрительной области), которые реагируют на углы определенных размеров и определенной ориентации в различных точках зрительного поля.
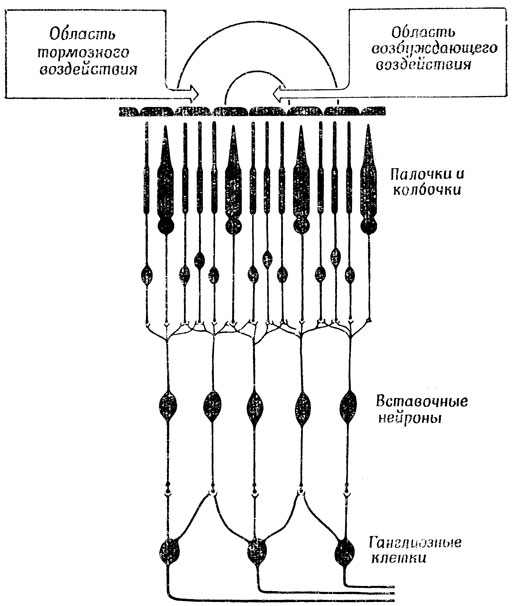
Рис. 29. Схема участка сетчатки (амакриновые и горизонтальные вставочные нейроны не изображены), на которой показано различное влияние палочек и колбочек на ганглиозные клетки: рецепторы, лежащие в центральной области, оказывают возбудительное действие, а лежащие на периферии - тормозное
Как видите, детекторы углов, существование которых мы предсказали из теоретических соображений (рис. 20), были действительно обнаружены в зрительной области коры головного мозга кошки [119] вместе с клетками усиления контраста [144], а латеральное коленчатое тело, по-видимому, обеспечивает предварительную обработку зрительной информации для выделения контуров. В зрительной системе лягушки детекторы угла обнаружены не были. Почему же для лягушек наше "предсказание" не оправдалось? Правдоподобным представляется следующее объяснение. У лягушки очень простое зрительно обусловленное моторное поведение: она не реагирует на зрительные стимулы, если не считать тех, которые вызывают у нее желание схватить или приготовиться к нападению (например, движущийся объект, напоминающий муху) или стремление спастись бегством (например, большой движущийся предмет). Короче говоря, она реагирует на локальные черты внешнего мира и не нуждается в информации, собранной с большего участка зрительного поля, для того чтобы направлять свое поведение. Таким образом, в ситуациях, в которых другие животные, например кошка, могут воспользоваться информацией об углах для того, чтобы свести информацию о контурах до необходимого минимума, так что лишь на высших уровнях интеграции возникает деенаправленное представление, основанное на "глобальных" свойствах зрительных стимулов, лягушке для того, чтобы действовать, достаточно "локальных" свойств (хотя для того, чтобы решить, на каком локальном свойстве основывать свое действие, лягушке понадобятся глобальные вычисления; см. разд. 7.2); таким образом, уместно допустить, что выход системы предварительной обработки зрительных образов у лягушки закодирован в терминах возможных действий и что одну группу клеток в сетчатке лягушки правомерно называть "детектором насекомых". Мы предполагаем, что управление движением глаз у млекопитающих весьма сходно с управлением движением всего тела у лягушки.
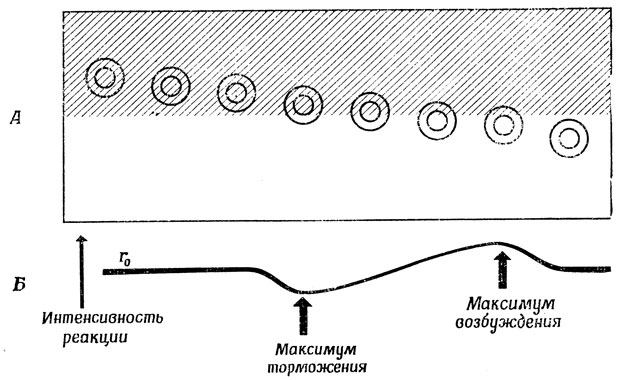
Рис. 30. Каждый из детекторов контраста с on-центром активизируется пропорционально разности стимуляции центральной области и периферии. Такие детекторы подчеркивают границу 'дублетом' отклонений от уровня покоя (сначала в одну, а потом в другую сторону). Клетки максимально возбуждены, когда они только что вошли в освещенную зону, и максимально заторможены перед самым выходом из нее
Обратите внимание, что тем самым мы утверждаем, что если предварительная обработка зрительных стимулов в ганглиозных клетках лягушки уже "ориентирована на действие", то у кошки даже такие "поздние" стадии предварительной обработки, как выделение углов в полях 18 и 19 коры головного мозга, "нейтральны по отношению к действию" и основаны лишь на том, что закономерности окружающей среды дают возможность хорошо описывать объекты с помощью точек максимальной кривизны на их контурах. Отложим обсуждение "распознавания образов" более высокого уровня до разд. 4.3, а здесь лишь упомянем работу Арбиба [8], посвященную проблемам сложности и эволюции, в которой высказываются предположения о том, каким образом различные функции сети могут благоприятствовать различным способам кодирования информации. Это может служить еще одним аргументом в пользу нашего предположения о том, что "деенаправленное кодирование" играет важную роль в системах восприятия.
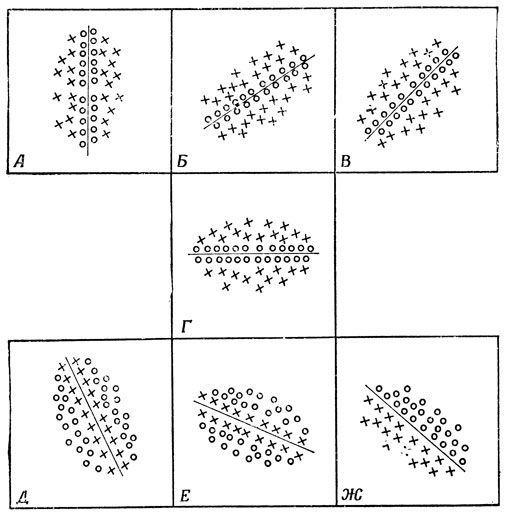
Рис. 31. Хьюбел и Визель обнаружили в коре головного мозга кошки популяцию клеток, которые они назвали простыми; в рецептивных полях этих клеток имеются on- и off-области (разделены прямой линией). Эта граница располагается по-разному в разных клетках, а диаметр всего рецептивного поля таких клеток кошки соответствует зрительному углу примерно 4°. Кружками показаны возбудительные (on) участки, а крестиками - тормозные (off)
Заложив некоторые основы для нашей гипотезы о деенаправленности восприятия и получив некоторые начальные сведения о нейронах и о строении нервной системы, мы отложим пока проблемы строения и функции мозга и займемся в двух следующих главах тем вкладом, который могут сделать теория систем и исследования по искусственному интеллекту в изучение мышления и поведения.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://informaticslib.ru/ 'Библиотека по информатике'